


 Test acouphènes
Test acouphènes
 Test auditif
Test auditif
L’oreille perçoit des ondes sonores qu’elle code puis interprète à travers un réseau structurel et neuronal complexe. Il faut donc avant tout définir les principaux fondements du son afin de comprendre le fonctionnement de l’oreille.
-
 Être rappelé
Être rappelé
-
Trouver un centre
-
Demander un rendez-vous
Être rappelé
Demandez un rendez-vous
"*" indicates required fields
Être rappelé
Rappel sur le son et la psychoacoustique
L’onde sonore
C’est la propagation de perturbations mécaniques dans un milieu élastique. Le son est produit sous forme de vibration mécanique d’une matière (solide, liquide, gazeux), qui se déplace sous forme d’ondes longitudinales, sans transport de matière. C’est cette variation de pression qui pourra être perçue par l’oreille, en fonction de sa fréquence et de son intensité.
Dans l’air par exemple, la source sonore met en vibration des particules d’air, qui ne se déplaceront que de quelques micromètres, entraînant un déplacement des particules d’air proches avant de revenir à leur position normale et ainsi de suite sous forme d’une chaîne de transmission de la perturbation.
L’onde sinusoïdale créée est longitudinale dans la plupart du temps (les molécules se déplacent parallèlement au sens de propagation de l’onde) et sphérique (se propage dans toutes les directions). A distance de la source, cette onde sonore tend vers une onde plane. Dans les solides, en raison des contraintes du matériau, l’onde sonore se propage de manière beaucoup plus complexe et peut être transversale.
Constantes
On définit la période T en secondes, comme l’intervalle de temps entre deux états vibratoires identiques et successifs d’un point du milieu dans lequel l’onde se propage. La fréquence, en Hertz, est définie comme l’inverse de la période. Elle est également définie par son intensité, correspondant à l’énergie qui traverse chaque seconde une surface unitaire perpendiculaire à la direction des ondes sonores (importance du déplacement des molécules par rapport au point de repos).
- La durée d’un son est le temps pendant lequel le milieu est perturbé.
- La vitesse de propagation d’une onde sonore (ou célérité), dépend des caractéristiques du milieu : sa compressibilité et sa masse volumique. Ces dernières sont influencées par la température (qui augmente la célérité) et la pression (qui diminue la célérité). Dans tout milieu, le son subit une résistance à sa transmission appelée impédance acoustique. Lors d’un passage du son d’un milieu A à un milieu B, une partie de l’onde sera transmise, et une autre sera réfléchie, réfractée, difractée ou absorbée.
Les différents types de sons
Vous comprendrez bien que l’onde sonore perçue par l’oreille n’est jamais simple. On peut donc définir différents types de sons :
- Le son pur est idéal et n’existe pas dans la nature. La vibration sonore est réduite à une seule fréquence. Il est utilisé pour l’audiométrie clinique, souvent en pulsé (interrompu périodiquement pour une meilleure perception).
- Le tone burst est un son pur émis pendant quelques millisecondes.
- Le clic est une impulsion rectangulaire émise pendant 100ms. Celui-ci est très utilisé en audiométrie objective.
- Un son pur modulé est un son dont l’amplitude ou la fréquence varie de façon périodique.
- Le son complexe est un mélange de sons purs.
- Le speech noise est un son correspondant au spectre de la voix parlée, il est utilisé pour le masquage en audiométrie vocale.
- Le bruit est une émission de sons aléatoires dont le spectre sonore est continu. Il peut être stationnaire (émis en permanence) ou transitoire. Le bruit en audiologie peut être tout type de son perçu comme un son non désiré et dérangeant.
- Le bruit blanc est un son dans lequel la densité spectrale de puissance est la même pour toutes les fréquences de la bande passante.
- Le bruit bande étroite est un bruit blanc auquel on applique un filtre passe bande. Il est donc défini uniquement sur une bande de fréquences choisie. Il est très utilisé pour le masquage en audiométrie tonale ou pour la prise en charge des acouphènes.
- Le bruit rose est le même principe qu’un bruit blanc mais avec une densité spectrale identique par bande de fréquences. Il possède donc un spectre constant sur une échelle logarithmique et se rapproche de la sensibilité de l’oreille humaine.
La représentation du son
Le son peut être représenté de différentes manières selon ce que l’on souhaite observer.
La représentation temporelle montre l’intensité du signal sonore en fonction du temps. Il permet d’observer l’enveloppe temporelle du signal. Cette enveloppe temporelle est très importante pour la perception de la parole dans le silence.
La représentation fréquentielle permet d’observer la structure fréquentielle d’un signal sonore, montrant ainsi pour chaque fréquence, l’intensité du signal sonore. Cette structure fréquentielle est notamment très importante pour la perception de la musique.
Une représentation sous forme de spectrogramme ou sonagramme permet d’observer la composition fréquentielle d’un signal sonore en fonction du temps. L’intensité de chaque fréquence à un instant T est représentée sous forme d’un code couleur (bleu à rouge), permettant d’apprécier la répartition énergétique du son. Cela est utilisé pour analyser une parole par exemple et observer ainsi la composition d’une syllabe avec le fondamental laryngé, les harmoniques et les transitions formantiques.
Les mesures sont souvent réalisées à l’aide d’un sonomètre permettant une analyse en fréquences et en intensité.
Les décibels
L’unité de mesure de la pression acoustique est le Pascal. L’oreille humaine n’est pas sensible à toutes les pressions acoustiques. Par exemple, à 1000 Hz, l’intensité la plus faible perçue est de 2/100 000 Pa, tandis qu’au delà de 20 Pa, le son devient douloureux pour l’oreille. Cet écart de 1 million entre les deux n’est évidemment pas pratique à utiliser. Il a donc été convenu de transformer cette échelle en décibels, en passant sur une échelle logarithmique, avec la formule suivante : Lp= 10 log (p2/ pO 2), avec pO= 10-5 Pa
(Pression acoustique correspondant au seuil d’audition normal à 1000 Hz)
Le dB SPL (sound pressure level) :
Le dB SPL en découle et est l’unité de référence de l’intensité d’un son en audiologie. Le seuil juste audible à 1000 Hz sera ainsi de 0 dB SPL et le seuil de douleur de 120 dB SPL. La quantité 1 dB représente à peu près un écart minimum correctement perçu par l’oreille humaine. L’intensité augmente avec la pression acoustique au carré. Si la pression acoustique est doublée, le niveau sonore augmente de 6 dB.
L’addition de plusieurs sons de sources différentes ne donne pas en revanche une addition simple du niveau sonore. Par exemple, si deux sources sonores différentes produisent un niveau identique, le niveau qui en résulte sera de 3 dB en plus. Pour 10 sources, il sera de 10 dB en plus. Mais cela n’est pas si simple à calculer en champs diffus lorsque le son se réfléchit sur les parois et que des interférences se produisent en créant d’autres ondes stationnaires.
Le dB HL (hearing loss) :
Le dB HL a été créé pour répondre à la sensibilité différente de l’oreille selon les fréquences. En effet, pour chaque fréquence, le minimum d’intensité perçu sera différent en dB SPL. Une conversion permet de ramener tous les seuils de perception (minimum d’intensité audible) à 0 dB HL pour une meilleure visibilité des graphes. Cette conversion a été faite d’après des valeurs mesurées sur une population de normo-entendants de 20 ans. Cela permet de tracer un audiogramme représentant les seuils d’audition du patient testé par rapport à la référence 0 dB HL et de mesurer sa perte auditive. Le Décibel HL est l’unité de mesure utilisée en audiométrie clinique et sert à quantifier la capacité auditive sur un graphique appelé audiogramme.
Les autres Décibels :
Une pondération peut être appliquée selon le sujet étudié. Par exemple, le Décibel A (dB A) permet de mieux prendre en compte les dangers potentiels d’un son perçu en moyenne sur une journée. Il consiste à corriger l’intensité de chaque fréquence en fonction de la sensibilité relative de l’oreille pour des bruits de niveau moyen. Le Décibel C (dB C) permet de mettre en évidence la dangerosité des pics d’intensité. En pratique, ils sont très utilisés en médecine du travail pour mesurer le risque potentiel d’un environnement sonore et l’adaptation de poste nécessaire ou encore pour mesurer les niveaux de bruit résultant dans un appartement proche d’un environnement bruyant.
La sonie
C’est la perception auditive subjective de l’intensité acoustique d’un son. Elle augmente notamment lorsque la pression acoustique augmente et elle permet notamment de distinguer un son faible d’un son fort.
- Le seuil absolu est l’intensité minimale perçue par une personne à une fréquence donnée.
- Le seuil d’inconfort est mesuré sur des intensités fortes, il représente l’intensité minimale provoquant une sensation auditive inconfortable.
La différence entre le seuil absolu et le seuil d’inconfort mesurée à chaque fréquence permet de définir une dynamique auditive ou un champ auditif. Ce champ auditif représente la zone dans laquelle les sons seront codés par l’oreille humaine.
Le champ auditif complet de l’être humain est situé à peu près entre 20 et 20 000 Hz et correspond à 10 octaves environ. On peut considérer que le champ auditif humain utile ou champs dynamique, se situe entre 0 et 120 dB SPL pour des fréquences allant de 125 Hz à 8000 Hz. Au-delà de ces fréquences, l’oreille humaine ne perçoit plus aussi bien les sons et la dynamique est réduite.
Ces fréquences correspondent à une spécialisation de l’oreille humaine dans l’écoute des sons correspondant à la voix humaine, étant un être sociable et dont la survie dépend de la communication avec ses semblables. C’est le cas également pour d’autres espèces qui n’ont donc pas le même spectre fréquentiel que celui de l’Homme. On parle d’infrasons pour les fréquences inférieures à 20 Hz et d’ultrasons pour les fréquences supérieures à 20 000 Hz. La chauve-souris par exemple utilise les ultrasons pour s’orienter, tandis que l’éléphant utilise les infrasons pour communiquer à distance.
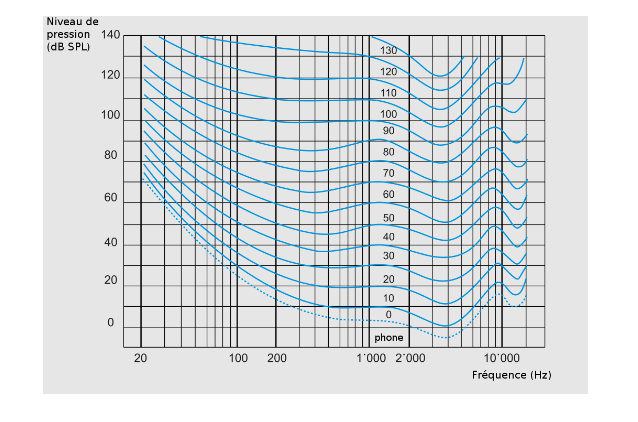
Les courbes isosoniques (Robinson et Dadson 1956)
Ce sont des lignes rassemblant les sons des différentes fréquences du champ auditif perçues à la même intensité. Elles ont été normées à l’aide de 50 normo-entendants en écoute binaurale champ libre. Ces courbes isosoniques sont exprimées en phone. Le phone est égal au dB SPL à 1000 Hz mais diffère aux autres fréquences pour lesquelles l’oreille n’a pas la même sensibilité.
Le sone découle du phone pour mettre en évidence l’évolution de la sensation auditive. La formule est la suivante : à partir de 40 phones, sone = 2 (phone-40) /10.
La sonie représente donc l’évolution de la sensation auditive sur des niveaux de pression sonore différents. On observe qu’elle croît à peu près comme le logarithme de la stimulation (Weber-Fechner, 1860). Elle croît également beaucoup plus rapidement sur les fréquences à l’extrémité du champ auditif où la dynamique est réduite. De même, en cas de perte auditive, on peut observer un phénomène de recrutement lorsque la sensation d’intensité augmente plus vite que le niveau acoustique.
Le seuil différentiel d’intensité représente la plus petite variation d’intensité créant une différence de sonie. Il est également variable selon la fréquence, l’intensité du son et selon la durée d’émission du signal. Il est de 2 dB au seuil d’audition et de 0,4 dB à 90 dB.
La tonie
La tonie est la sensation de hauteur subjective permettant de classer deux sons de fréquence différente (par exemple, un son grave et un son aigu). La perception du timbre est un phénomène complexe permettant d’identifier une source sonore par rapport à une autre (par exemple, deux instruments de musique différents ou deux voix différentes). Et ce, même si le son émis est de même fréquence et de même intensité. Il dépend notamment de la richesse spectrale et des harmoniques d’un son.
Les bandes critiques
La cochlée est constituée d’une série de bandes fréquentielles juxtaposées. Ces bandes fréquentielles ont été mises en évidence par des expériences psychoacoustique. Par exemple, deux sons de fréquence très proche ont été envoyés en même temps. Tant qu’ils restaient dans la même bande fréquentielle, la sonie restait identique et augmentait lorsque l’on dépassait la bande critique.
On peut les comparer à des filtres passe-bande centrés sur une fréquence. Il y en aurait 24, définies selon leur fréquence centrale et non figées. Cette sélectivité fréquentielle peut être mise en évidence par des courbes d’accord. Elles sont obtenues en émettant un son pur de 10 dB à une fréquence donnée et en relevant l’intensité minimale qu’il faut pour le masquer avec d’autres sons purs de fréquences différentes. On obtient alors une courbe formant un V, plus ou moins étroit selon la sélectivité de l’oreille à chaque fréquence.
Le seuil différentiel de hauteur est la plus petite variation sonore perceptible et dépend de la fréquence, de l’intensité et de la durée.

Effet de masquage
Lorsque plusieurs sons sont envoyés simultanément, ils sont perçus en général indépendamment. Mais il arrive qu’un son empêche la perception d’un autre son : on parle d’effet de masquage.
Celui-ci dépend de plusieurs facteurs physiologiques. Un bruit sera généralement plus masquant si la fréquence du son émis est proche du bruit et l’effet de masquage augmentera avec l’intensité. De plus, les basses fréquences masquent plus fortement les hautes fréquences. Le masquage apparait également au niveau central, un bruit masquant baissant la sonie perçue du son pur.
Audition binaurale
L’audition binaurale, contrairement au monaural, est le fait d’entendre avec les deux oreilles. Elle permet d’améliorer la sensation d’intensité de 3 dB au seuil et de 6 dB au-delà de 35 dB HL. Elle permet également une meilleure localisation des sons et une amélioration de la compréhension dans le bruit. En effet, le rapport signal sur bruit est amélioré par le démasquage binaural d’environ 10 dB. C’est la raison pour laquelle lorsqu’on perd l’audition d’une oreille (cophose unilatérale), nous devenons très gêné en milieu bruyant avec des difficultés importantes pour comprendre la parole. Une cophose unilatérale peut s’appareiller avec un système CROS.
Anatomie et fonctionnement de l’oreille
Un son est toujours produit par une vibration, comme celles d’un tambour ou des cordes vocales. Ces vibrations se propagent dans l’air sous la forme d’une onde. Pour rendre le son intelligible, l’Homme dispose d’un organe complexe et fragile : L’oreille.
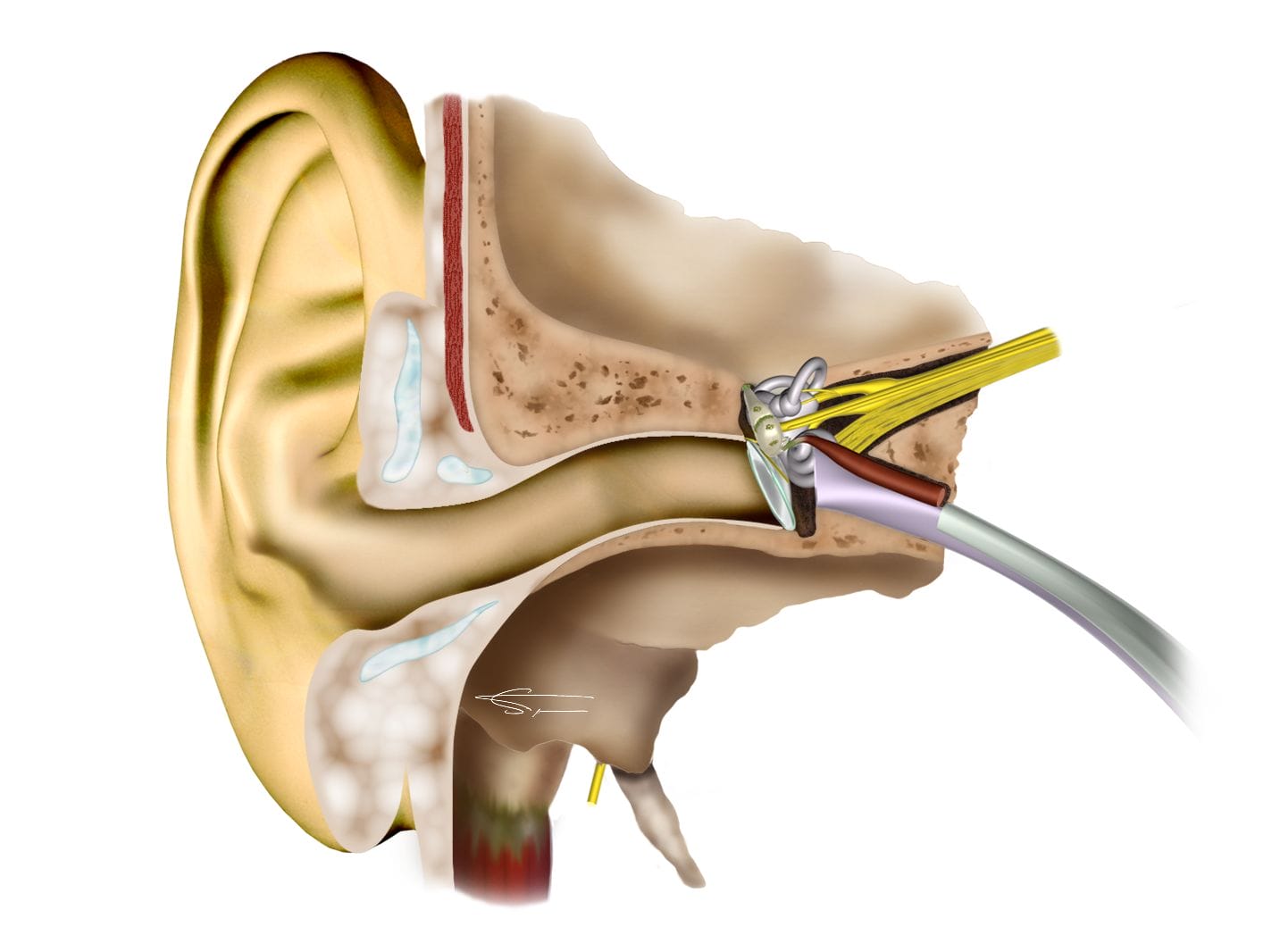
Fonctionnement de l’oreille
Les ondes sonores sont captées et concentrées vers le tympan par l’oreille externe à la manière d’une parabole. Le tympan transmet les vibrations aux osselets, puis à l’oreille interne. C’est là, dans la cochlée que se fait le véritable traitement des sons. Dans cette spirale osseuse remplie de liquide, des cellules ciliées vont capter les vibrations pour les transmettre au cerveau sous forme de micro signaux électriques via le nerf auditif.
Une perte d’audition survient lorsqu’une ou plusieurs parties de l’oreille sont déficientes. C’est le cas de 466 millions de personnes dans le monde.
Composition de l’oreille
L’oreille est l’organe sensoriel de l’audition. Elle est située dans un rayon de 4 cm, au niveau de l’os temporal. Cet os temporal se compose de plusieurs parties : une partie écailleuse contenant l’os zygomatique, une partie mastoïdienne, et une partie pétro-tympanique ou rocher. Le rocher est l’os principal qui abrite l’oreille. Il se compose d’une partie tympanique comprenant le conduit auditif externe, et d’une partie pétreuse comprenant l’oreille moyenne et interne. Le rocher accueille des structures nobles telles que la veine jugulaire et l’artère carotide interne.
Elle se compose de trois parties : l’oreille externe, qui réceptionne le son, l’oreille moyenne qui transporte le son vers l’oreille interne qui va le coder en signal électrique complexe.
L’oreille externe
C’est la partie de l’oreille qui capte les sons, elle comprend le pavillon et le conduit auditif externe.
Le pavillon
Il présente une forme parabolique ainsi que des circonvolutions (hélix, anthélix …). Cette forme permet de concentrer les ondes sonores vers le conduit auditif externe, de mieux capter les sons latéraux et antérieurs, d’améliorer la localisation des sons, et de renforcer le son de 3 à 6 dB.
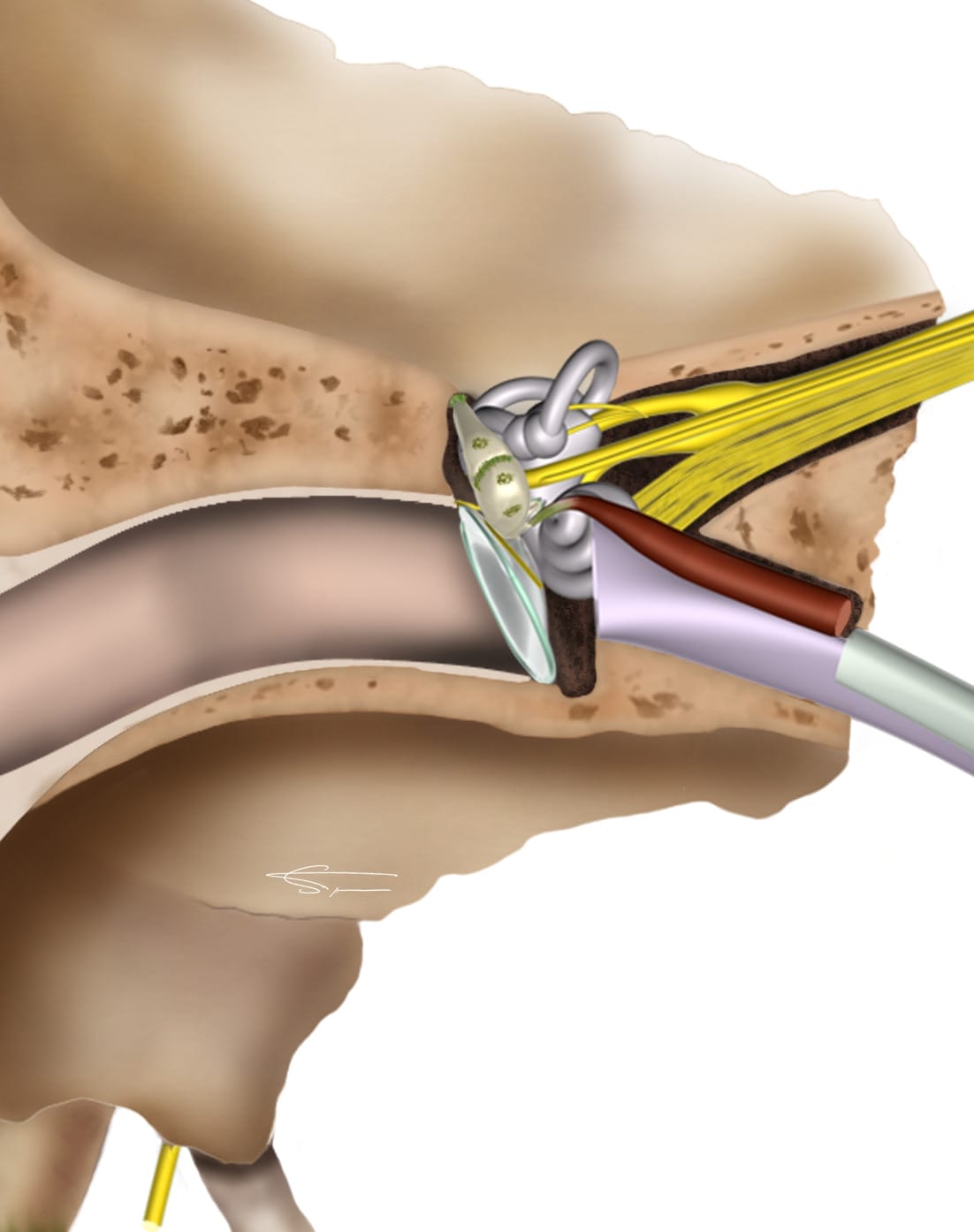
Le conduit auditif externe
D’environ 3 cm, il guide le son vers la membrane tympanique. Il présente deux courbures, et est formé d’un segment externe cartilagineux (recouvert d’une peau épaisse comportant des poils et des glandes cérumineuses à l’origine de la formation du cérumen), et d’un segment interne osseux (recouvert d’une fine couche cutanée dépourvue de follicules pileux et de glandes cérumineuses).
L’innervation sensitive de l’oreille interne est assurée par différents nerfs (X, V, VII bis, et C2/C3).
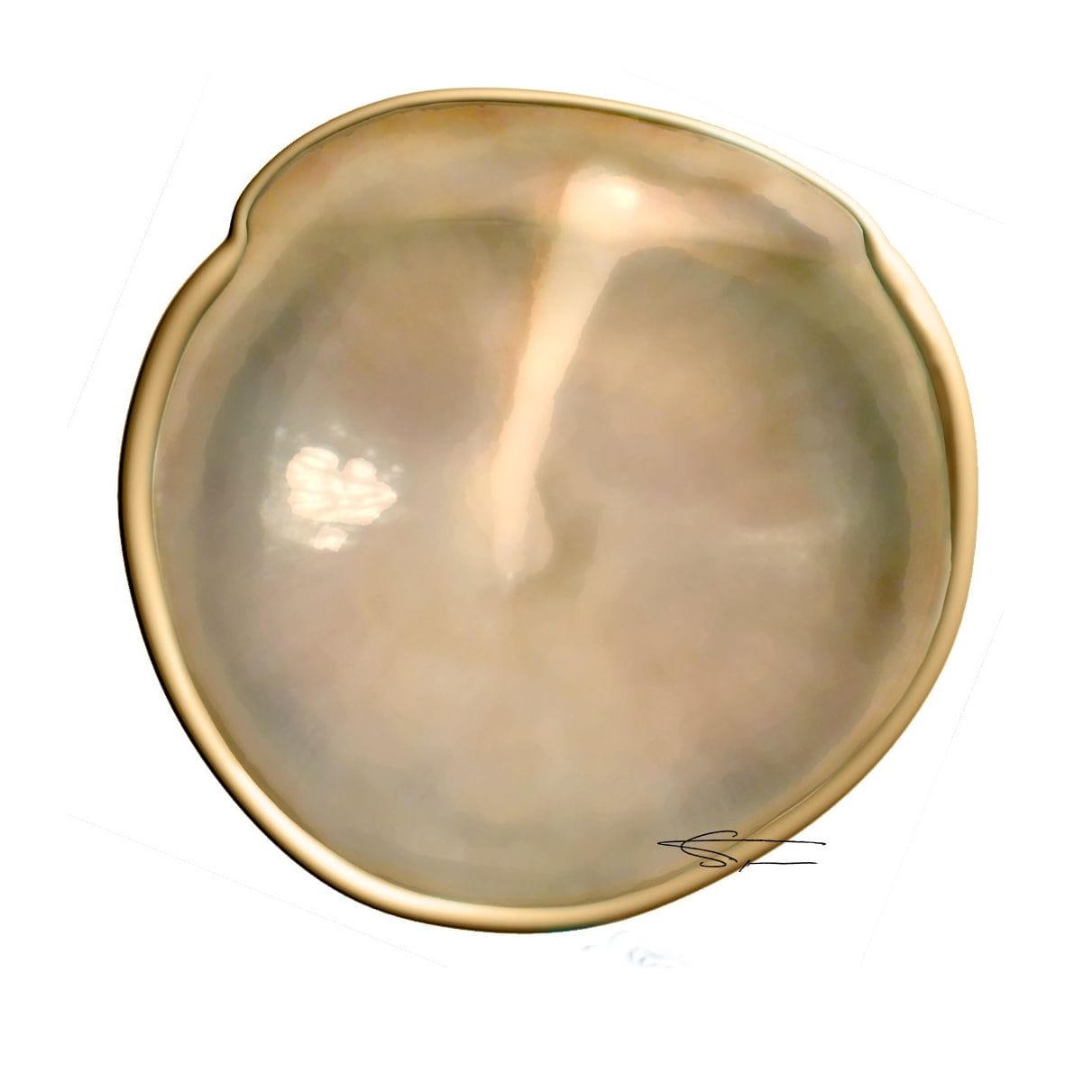
Le tympan
Le fond du conduit est fermé par le tympan, membrane vibrante constituée de deux parties :
- Majoritairement la pars tensa, constituée de 3 couches : la couche externe épidermique, la couche moyenne fibreuse, et la couche profonde muqueuse.
- La pars flacida ou membrane de Schrapnell, au-dessus des ligaments, très petite et plus fragile.
Le tympan s’insère dans le sillon tympanique par un anneau fibreux (Gerlach), et dans la courte apophyse du marteau par des ligaments tympano-malléolaires antérieurs et postérieurs au niveau de l’échancrure de Rivinus. Il présente sur sa face externe une dépression centrale (l’ombilic), en haut et en avant un point correspondant à la petite apophyse du marteau, en bas et en arrière la saillie du manche du marteau.
Un tympan fonctionnel est souvent révélé à l’otoscopie par un reflet de lumière appelé triangle lumineux de Politzer.
Fonctionnement
Les ondes sonores arrivant à l’oreille sont modifiées en ondes planes par les structures anatomiques de l’oreille. Ces dernières ont tendance à filtrer les basses fréquences et à renforcer les fréquences aigues.
Le conduit auditif permet de transmette le son de l’extérieur à l’oreille moyenne. Le pavillon permet de localiser les sons, tandis que la conque et le conduit auditif externe agissent comme un résonnateur naturel. Un gain de 10 dB est appliqué environ entre 1500 et 7000 Hz, avec un maximum vers 2800 Hz. Ce résonnateur varie selon la forme de l’oreille et sa taille (enfant vs adulte). La conque permet un gain à 5000 Hz, tandis que le conduit auditif externe apporte un gain supplémentaire à 3000 Hz. L’angle d’arrivée de l’onde à l’entrée du conduit auditif externe joue également un rôle dans la résonnance appliquée.
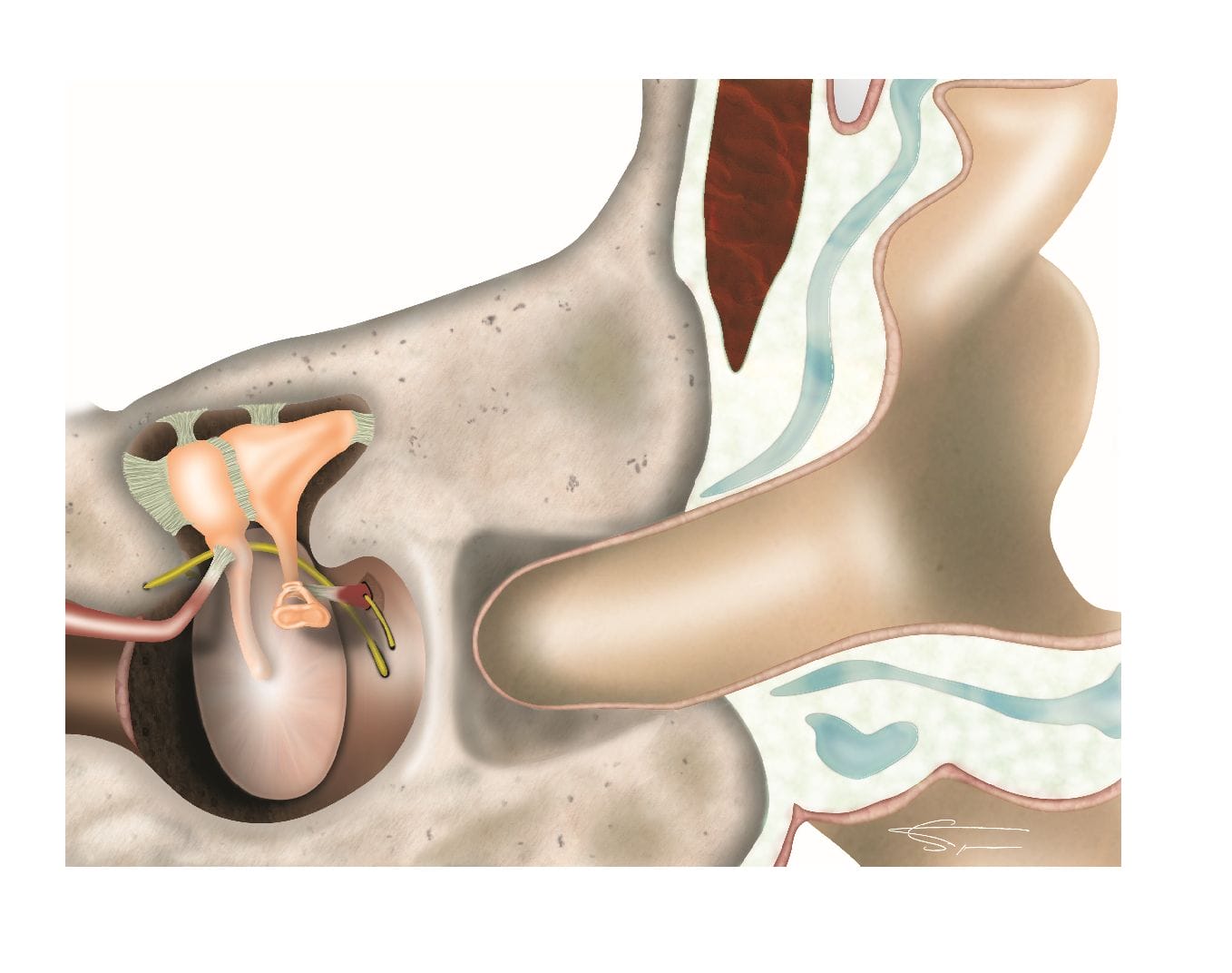
L’oreille moyenne
C’est une cavité cylindrique, délimitée par le tympan (au fond du conduit auditif externe), comprenant la chaîne des osselets (marteau, enclume et étrier) imbriqués les uns aux autres.
Elle est reliée au cavum (dans le nez) par la trompe d’eustache, et s’ouvre en arrière dans la mastoïde (antre mastoïdien). Elle contient des structures nobles telles que le nerf facial, et la corde du tympan. Cette corde du tympan naît du nerf facial, et contient des fibres du goût provenant du nerf intermédiaire de Wrisberg (VII bis).
L’oreille moyenne se divise en deux parties :
- La partie voutée, ou attique, située au-dessus de la membrane tympanique, comprenant les corps du marteau et de l’enclume, et les ligaments suspenseurs.
- La partie creuse, appelée atrium, contenant l’articulation incudo-stapédienne, et les fenêtre rondes et ovales.
Les parois de la caisse du tympan
- La paroi antérieure (ou haute), présente en haut de l’orifice du canal du muscle tenseur du marteau, en bas (partie antérieure haute de l’atrium) l’orifice de la trompe d’Eustache et l’orifice antérieur de la corde du tympan.
- La paroi inférieure, aussi appelé “plancher”, elle présente la région du golf de la jugulaire.
- La paroi postérieure, s’ouvre en haut sur la mastoïde par un orifice appelé l’aditus ad antrum, plus bas se trouve la pyramide qui donne issue au muscle de l’étrier et l’orifice postérieur de la corde du tympan.
- La paroi interne ou labyrinthique, sépare la caisse du tympan de l’oreille interne. Elle est limitée par canal de Fallope contenant le nerf facial. En haut, au niveau de l’attique, elle présente la saillie du canal semi circulaire externe, et en bas, au niveau de l’atrium la saillie du promontoire (ou tour basal de la cochlée). On observe également le sinus tympani contenant le muscle de l’étrier et le nerf de ce muscle. On peut également observer la fenêtre ovale, reliée à la platine de l’étrier, permettant la mise en vibration des liquides péri labyrinthiques de l’oreille interne. En dessous du promontoire apparaît également la fenêtre ronde compensant la pression liquidienne appliquée au niveau de la fenêtre ovale.
- La paroi externe ou tympanique est essentiellement formée de la membrane tympanique.
- La paroi supérieure ou crânienne est appelée le tegmen tympani.
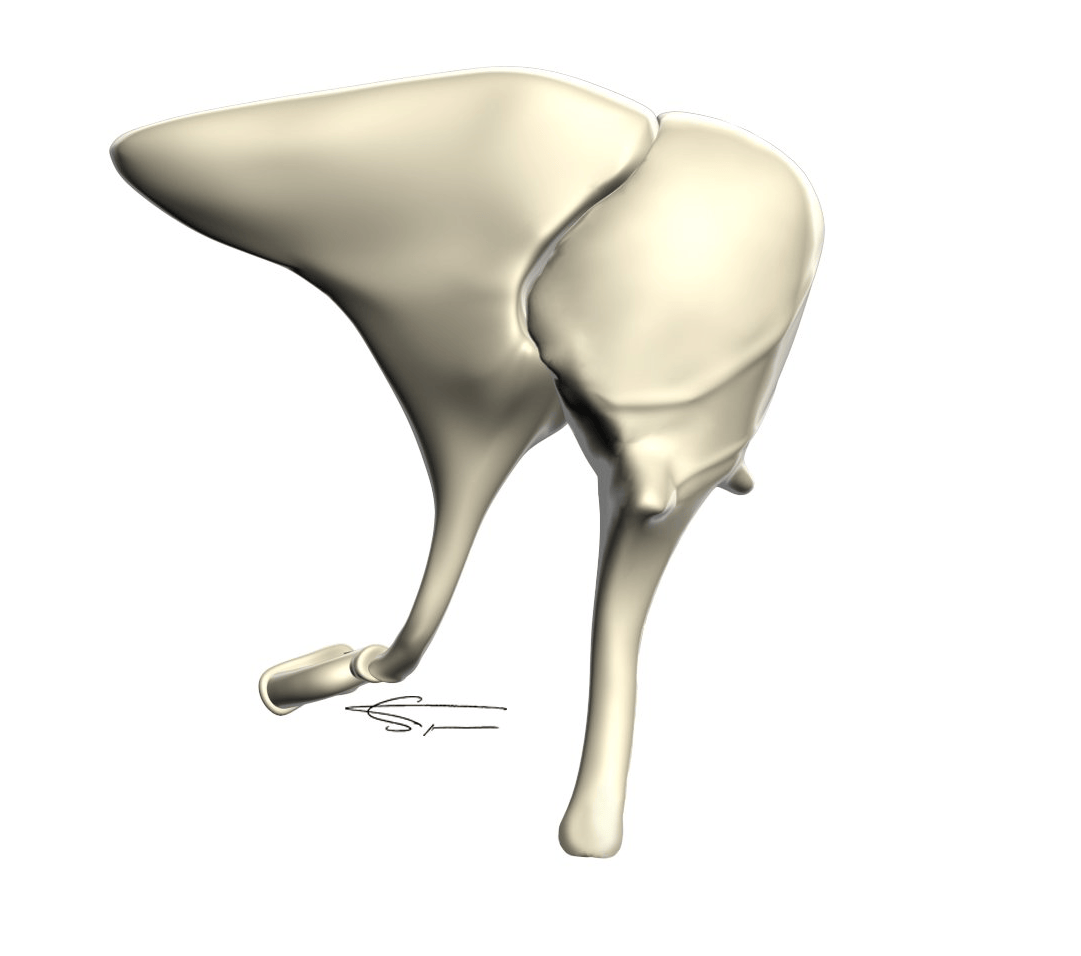
La chaîne des osselets
La chaîne ossiculaire comporte les trois plus petits os du corps humain, articulés entre eux dans la caisse tympanique. Elle permet la transmission du sons d’un milieu aérien (l’air) vers un milieu liquidien (l’oreille interne).
- Le marteau est le plus volumineux, avec un manche, un col et deux apophyses, ainsi qu’une tête articulée avec l’enclume. Il est relié au tympan par son manche, et est mis en vibration en premier.
- L’enclume en arrière du marteau, présente un corps, deux branches et une apophyse lenticulaire. Cet osselet augmente la vibration par un effet bras de levier créé par son anatomie.
- L’étrier comprend une tête, deux branches antérieures et postérieures et une platine. Cette platine met en vibration la fenêtre ovale.
Fonctionnement de la chaîne ossiculaire
Après la mise en vibration du tympan par les vibrations aériennes, l’oreille moyenne transforme ces dernières en variations de pression des liquides de l’oreille interne.
Elle agit même comme amplificateur naturel découlant de l’anatomie de la chaîne ossiculaire, afin d’éviter la perte d’énergie engendrée par le passage de l’air à l’eau : on parle d’adaptateur d’impédance.
L’adaptation d’impédance est la plus performante entre 800 et 1200 Hz. Aux basses fréquences la rigidité de la chaîne freine la vibration, tandis qu’aux hautes fréquences, c’est la masse de la chaîne qui la freine.
Le muscle du marteau, innervé par le nerf trijumeau (V moteur), et l’étrier, innervé par le nerf facial (VII), peuvent limiter la propagation du son. En effet, le nerf V et VII reçoivent des stimulations de l’olive protubérantielle, entrainant une contraction de ces muscles.
Le muscle de l’étrier permet de protéger naturellement l’oreille des sons forts à basse fréquence (de 70 à 100 dB HL), avec une atténuation maximale de 20 dB. Il agit en mettant en tension la membrane tympanique, diminuant ainsi la quantité d’énergie sonore parvenant à l’oreille interne. On parle du réflexe stapédien.
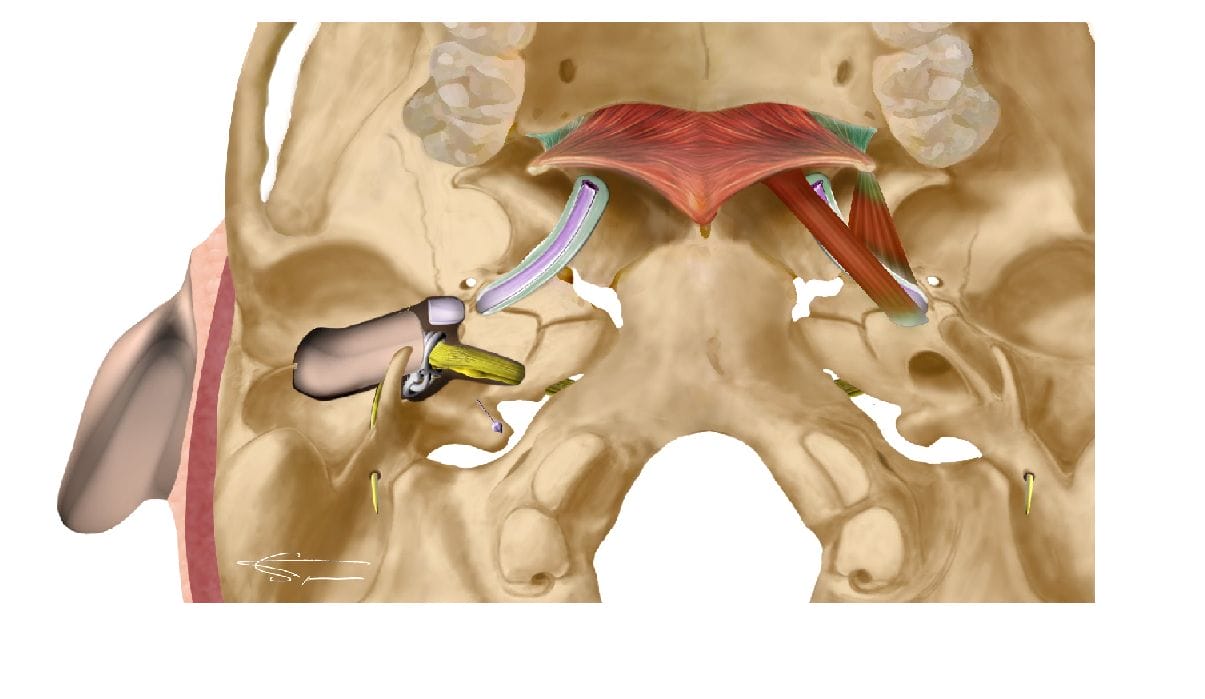
La trompe d’Eustache
Orientée en avant, en bas et en dedans, elle fait communiquer la caisse du tympan avec le rhino-pharynx. Elle est constituée d’une partie postérieure osseuse, et antérieure cartilagineuse. Elle contient dans son épaisseur des glandes pharyngées formant l’amygdale tubaire.
Un équilibre pressionnel parfait est essentiel pour le bon fonctionnement de la chaîne ossiculaire. Pour cela, des échanges gazeux sont réalisés au niveau de la muqueuse.
La trompe d’eustache permet également d’ajuster la pression de l’oreille moyenne, de manière rapide et régulière. Pour cela, elle s’ouvre et se referme brièvement, plus de 1000 fois par jour. Ce réflexe est permis par les muscles tenseurs et élévateurs du voile du palais, et est la plupart du temps réalisé lors de la déglutition, l’ouverture de la bouche, ou le bâillement. Il est possible d’équilibrer volontairement la pression entre l’oreille externe et l’oreille moyenne grâce à la manœuvre de Valsalva, du nom de son inventeur Antonio Maria Valsalva. Cette manœuvre est utile pour soulager l’oreille lors de variations de pression en montagne, en avion ou encore en plongée.
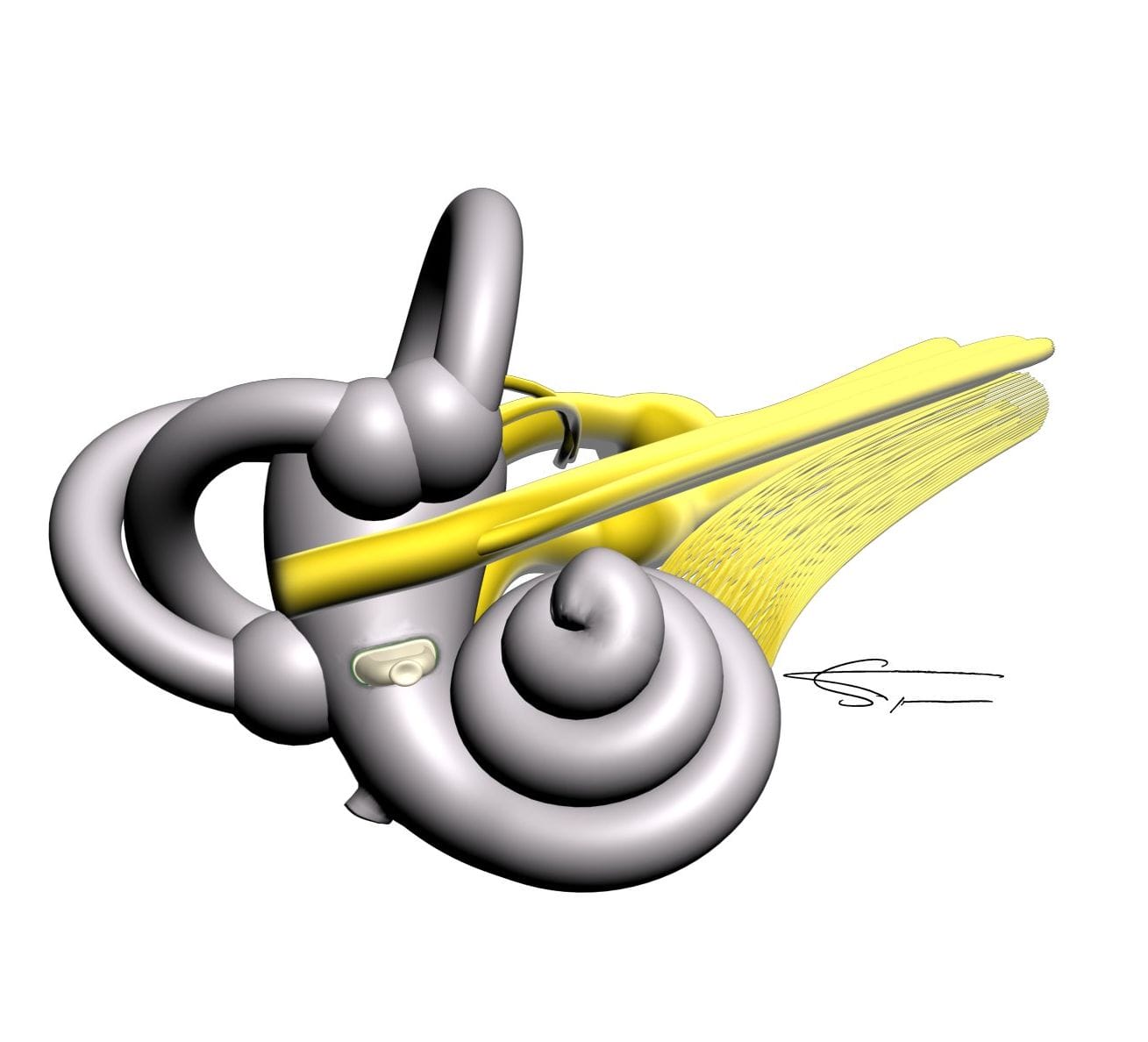
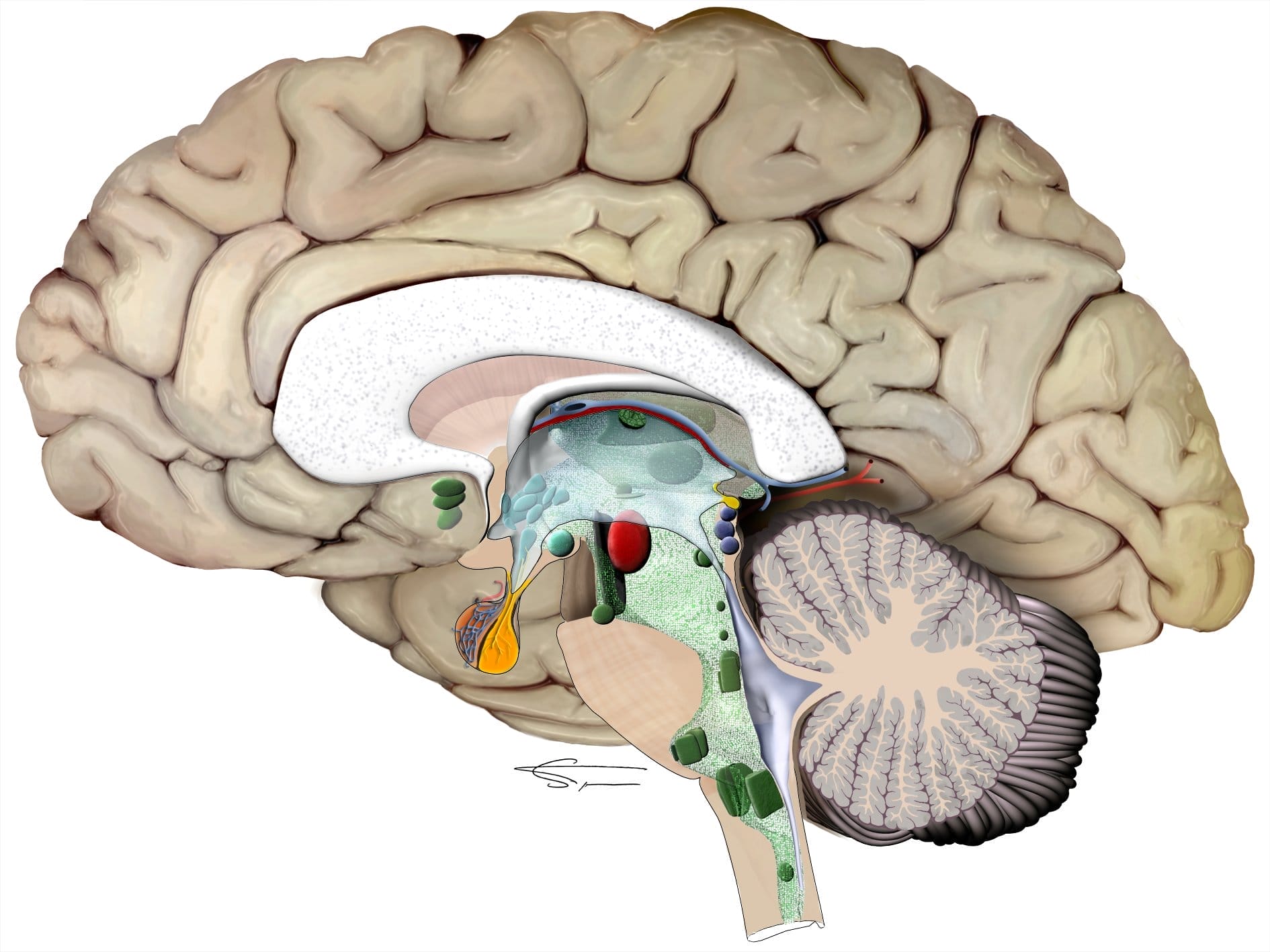
L’oreille interne
L’oreille interne contient le labyrinthe osseux, dans lequel se trouve le labyrinthe membraneux. Celui-ci contient les organes sensoriels sensibles au son et à l’équilibre. Il permet de transformer les informations mécaniques en influx nerveux qui seront conduits jusqu’au cerveau par des relais neuronaux complexes. Il se divise en deux parties : la cochlée, organe de l’audition, et le vestibule, organe de l’équilibre.
La cochlée est divisée en trois canaux, la rampe tympanique, la rampe vestibulaire et le canal cochléaire. Ce dernier contient l’organe de corti, possédant les structures cochléaires essentielles, telles que les cellules ciliées externes et internes, et étant le siège de la transduction mécano électrique.
Située dans la pyramide pétreuse, l’oreille interne permet donc de transformer les vibrations sonores en influx nerveux:
- le labyrinthe antérieur ou cochlée spécialisé dans l’audition transformant les vibrations acoustiques en influx nerveux.
- le labyrinthe postérieur ou vestibule spécialisé dans l’équilibre, comprenant l’utricule, le saccule et les trois canaux semi-circulaires. Elle a pour rôle de coder les accélérations rotatoires, horizontales ou verticales.
Vascularisation de l’oreille interne
Elle se fait à partir de l’artère basilaire ou du tronc basilaire qui donne l’artère cérébelleuse antéro-inférieure qui va donner naissance à l’artère labyrinthique (artère auditive interne) dans le conduit auditif interne .
L’artère labyrinthique va se diviser en deux parties :
- L’artère vestibulaire antérieure : elle vascularise la macule utriculaire, les ampoules et les parois membraneuses des c. s. c. supérieur et externe.
- L’artère cochléaire commune qui se divise en deux : l’artère cochléaire propre au canal cochléaire ; et l’artère cochléo-vestibulaire pour la partie initiale de la cochlée, l’ampoule et les parois membraneuses du c. s. c. postérieur ainsi que pour la macule sacculaire.
C’est une vascularisation sans suppléance donc toute souffrance vasculaire peut entrainer une dégradation plus ou moins importante de l’audition. C’est la cause de certaines surdités brusques ainsi que de surdités congénitales lors de souffrances fœtales (manque d’oxygénation du fœtus).

Composition de l’oreille interne
Le Labyrinthe osseux
L’oreille interne est formée du labyrinthe osseux, constitué d’un ensemble de cavités de formes complexes en communication les unes avec les autres, creusées à l’intérieur de la pyramide pétreuse du temporal.
Le labyrinthe antérieur osseux s’enroule sur deux tours et demi de spire autour du modiolus (accueillant le paquet vasculo-nerveux cochléaire). On peut observer sur le labyrinthe la fenêtre ovale et ronde.

Le Labyrinthe membraneux
Le labyrinthe membraneux est contenu dans le labyrinthe osseux qui le protège et auquel il est attaché par du tissu conjonctif.
Il est constitué de trois tubes, la rampe vestibulaire et la rampe tympanique, séparées par la lame spirale, qui, à sa partie distale, se dédouble en un canal triangulaire appelé le canal cochléaire. Ce dernier contient l’organe de corti, l’organe sensoriel de l’audition.

La rampe tympanique et la rampe vestibulaire
La rampe tympanique et la rampe vestibulaire sont remplies d’un liquide comparable au liquide céphalo-rachidien appelé périlymphe, riche en sodium (140 mmol/L) et en chlorure, et pauvre en potassium K+ (3 mmol/L)).
Elles communiquent entre elles par un petit orifice situé à l’apex de la cochlée nommé hélicotréma.
Lors de la mise en vibration de l’étrier (3ème os de la chaîne ossiculaire), la fenêtre ovale est mise en vibration, mettant en vibration les liquides dans la rampe vestibulaire (ou supérieure). La vibration de ces liquides incompressibles est transmise dans la rampe tympanique (ou inférieure) et déplacent la fenêtre ronde.
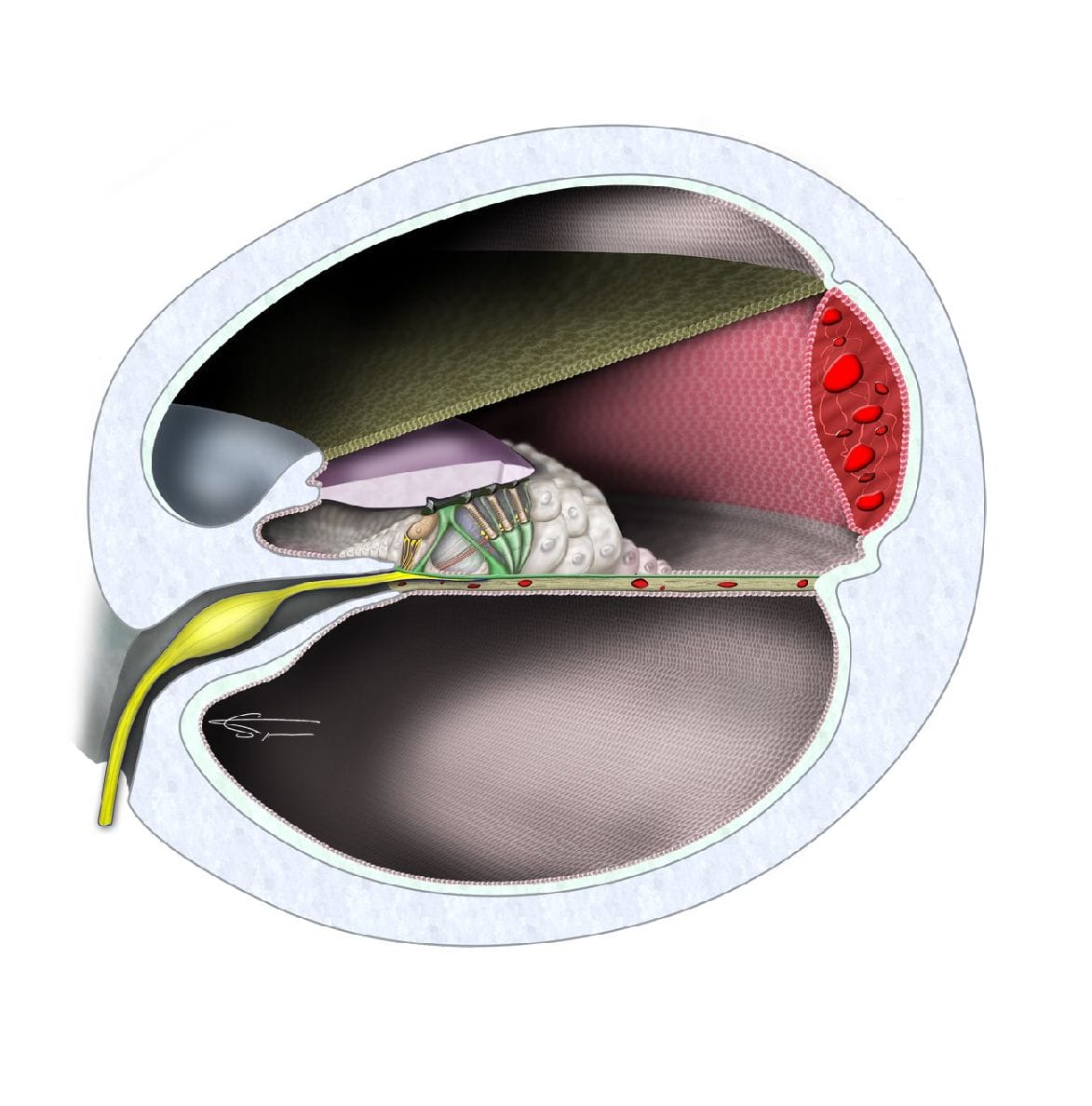
Le canal cochléaire
C’est le labyrinthe membraneux qui est le siège des récepteurs sensoriels. De lui naissent les grandes voies nerveuses acoustiques et vestibulaires qui se réunissent pour former le nerf auditif (VIII), qui emprunte le conduit auditif interne pour gagner la fosse cérébrale postérieure et le tronc cérébral.
Le canal cochléaire est délimité au-dessus par la membrane de Reissner. Sa paroi inférieure est la membrane basilaire sur laquelle repose l’organe de Corti. Ce dernier est l’organe sensoriel de l’oreille interne : il permet la transduction mécano-électrique du son (passage de vibrations mécaniques à un influx nerveux).
Les fibres du nerf cochléaire naissent de l’organe de corti, et s’engagent dans le canal de la lame spirale osseuse pour former le ganglion spiral de Corti (possédant le corps cellulaire du nerf cochléaire).
L’endolymphe
Le canal cochléaire contient du liquide endolymphatique, riche en potassium K+ (150 mmol/L) et en chlorure, et pauvre en sodium (1 mmol/L). Il est isolé du labyrinthe osseux par la périlymphe. L’endolymphe est produite par la strie vasculaire. Cette paroi externe du canal cochléaire est très riche en vaisseaux sanguins et permet de produire du potassium K+.
Le système endolymphatique est composé du canal endolymphatique qui naît de la réunion des canaux utriculaire et sacculaire, pénétrant dans l’aqueduc du vestibule osseux et reliant le vestibule à l’endocrâne. Il est également composé du sac endolymphatique situé dans la partie distale du canal endolymphatique. Une partie se trouve dans l’aqueduc du vestibule, l’autre dans un dédoublement de la dure-mère, sur la face postéro-supérieure du rocher. Le sac endolymphatique a un rôle important dans le maintien de l’homéostasie des liquides labyrinthiques. Il permet notamment de stocker et résorber l’endolymphe.
Composition ionique (mM)
La périlymphe Na+ 154, K+ 3, Cl- 128
L’endolymphe Na+ 1, K+ 161, Cl- 131
Le potentiel endocochléaire
Un potentiel négatif de -80 mV est créé entre la sécrétion active de K+ par la strie vasculaire dans l’endolymphe (120 mV), et la cellule ciliée (40 mV). Un transport d’ions K+ s’effectue alors dans la cellule, de manière passive et sans dépense d’ATP.
Ce potentiel endocochléaire est le moteur de l’oreille interne. Cela est permis par la loi de diffusion passive des ions K+, du milieu le plus concentré en ions au milieu le moins concentré : de l’endolymphe à la cellule ciliée, puis du corps cellulaire de la cellule ciliée, à la périlymphe.
Le cycle du potassium
Le potassium est recyclé par un processus complexe appelé « cycle du potassium ». Il passe des fibrocytes du ligament spiral aux cellules basales, puis des cellules basales aux cellules intermédiaires, par des jonctions communicantes, composées de connexons.
Ces connexons sont composés de 6 connexines (protéines transmembranaires), disposées sous forme d’hexamère créant un canal. Les connexines 26 et 30 notamment sont fortement exprimées dans le mur latéral et la strie vasculaire. Leur mutation entraîne des surdités, la plus répandue étant la mutation du gène GJB2 codant la connexine 26. Cette mutation entraine une surdité bilatérale congénitale isolée (non syndromique) de type DFNB1.
D’autres molécules sont importantes pour transporter et réguler le taux de potassium :
Le canal potassique Kir4.1 (gène KCNJ10), qui permet de faire sortir le potassium des cellules intermédiaires à l’espace intrastrial, la Na, K-ATPase qui régule la concentration de potassium dans cet espace, et, avec le co-transporteur NKCC1, l’entrée de potassium dans la cellule marginale.
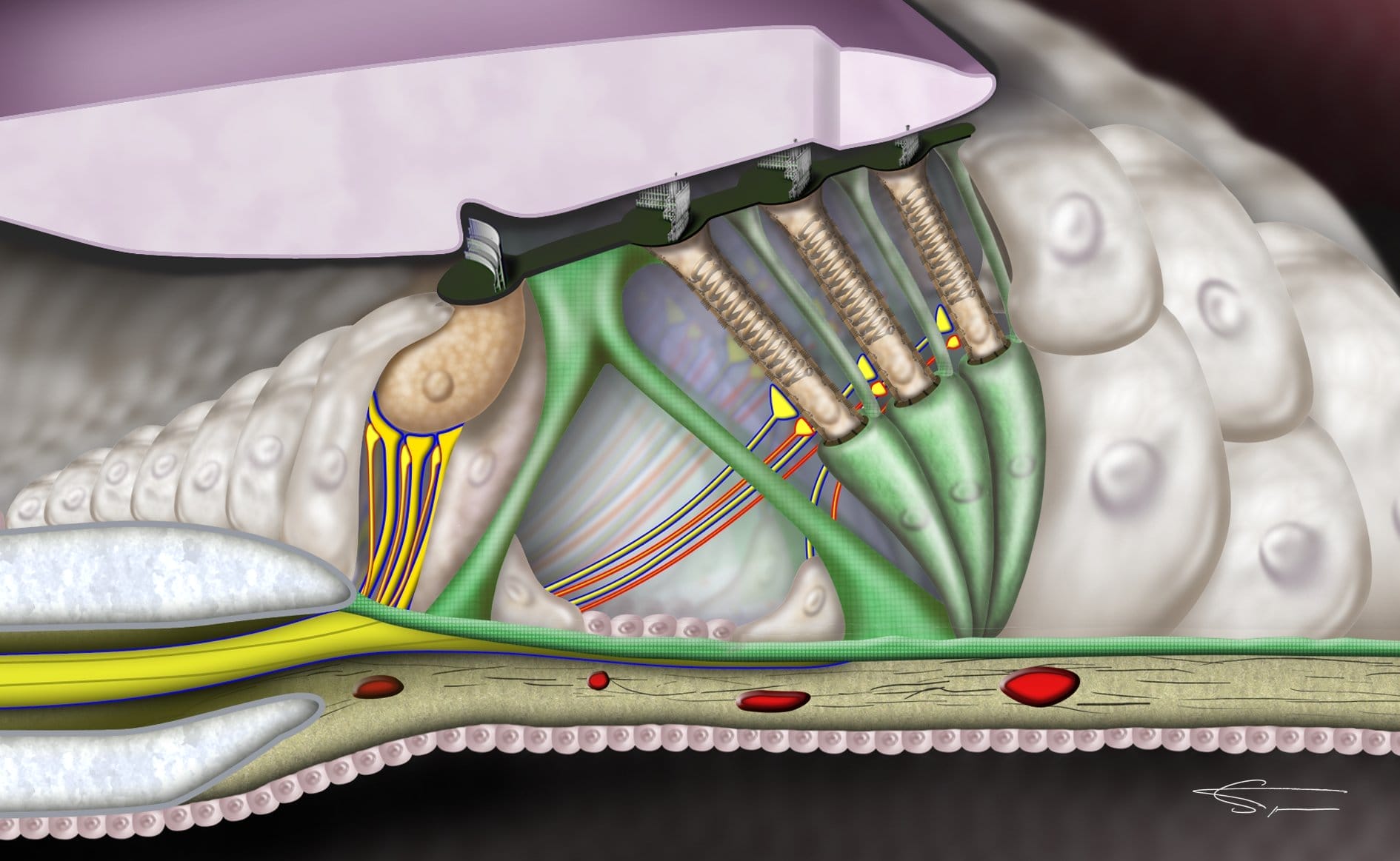
L’organe de corti
L’organe de corti est un mécanorécepteur. Il contient des cellules sensorielles appelées cellules ciliées (car elles sont coiffées de cils). Parmi elles, on distingue les cellules ciliées internes, et les cellules ciliées externes.
Les cellules ciliées externes et internes sont séparées par des piliers (pilier interne et pilier externe), au centre desquels on trouve le tunnel de Corti contenant de la perilymphe. Elles sont entourées et maintenues par des cellules de soutien (cellules de Deiters et de Hensen).
Au-dessus des cellules ciliées, on peut observer la membrane tectoriale, une structure épaisse recouvrant tout l’organe de Corti. Elle est solidement attachée aux cellules ciliées externes par la cuticule (ou plaque cuticulaire), dans lesquelles sont implantés les stéréocils.
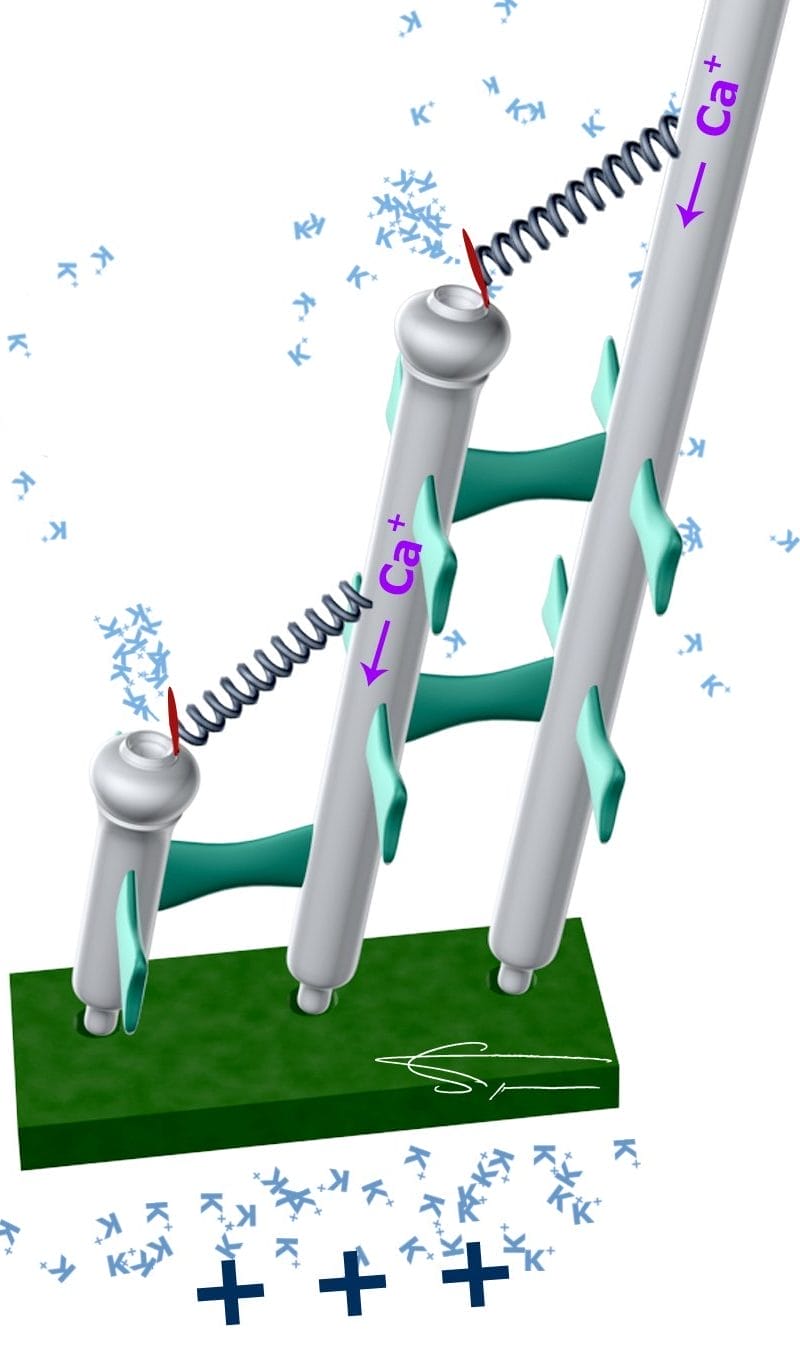
La transduction mécano-électrique au sein de l’organe de corti
Les cellules ciliées possèdent des cils ou stéréocils (environ 100) répartis sur 3 rangées, des plus grands aux plus petits, reliés entre eux par des tip-links à l’extrémité. Ces cils sont notamment constitués de filaments d’actine. Les cils baignent dans l’endolymphe tandis que le reste de la cellule baigne dans la périlymphe. Ces cils, voltage dépendants, sont très importants dans la transmission de l’information sonore : ils sont le siège de la transduction mécano-électrique.
Lors de l’arrivée de l’onde sonore, les longs cils sont souvent les premiers défléchis, entraînant la déflexion des deux autres cils plus courts. Ces derniers possèdent des canaux ioniques sensibles à l’étirement. Ces canaux cationiques, localisés à la base des tip-links (2 canaux par tip links), s’ouvrent, entraînant l’entrée de K+ dans la cellule ciliée qui se dépolarise.
Le cation Ca2+ entre également dans la cellule. Puis les canaux K+ se referment rapidement, prêts pour une autre stimulation. Cette fermeture, appelée mécanisme d’adaptation, possède une composante rapide (liée à la fixation du calcium sur les canaux), et une composante lente (liée à l’activation par le calcium du déplacement de la myosine le long des filaments d’actine des stéréocils).
Les stéréocils permettent notamment pour la cellule ciliée externe, une augmentation de sa sensibilité et de sa spécificité.
Les cellules ciliées possèdent des particularités physiologiques propres, avec un seuil d’activation (rhéobase), un temps de latence, une période réfractaire, une limite, conditionnant la transduction mécano-électrique du son.
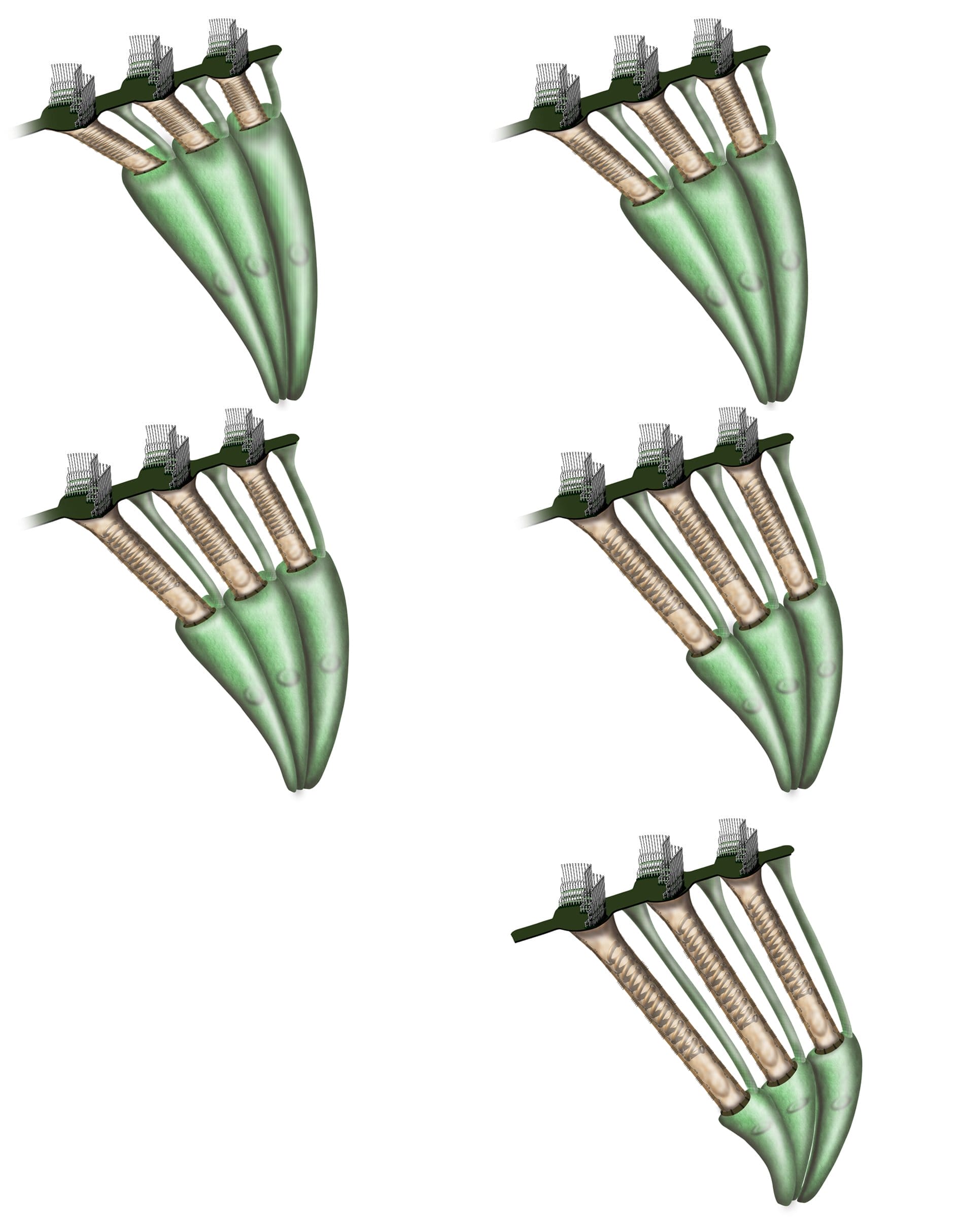
Les cellules ciliées externes
Les cellules ciliées externes (CCE), de forme cylindrique, sont organisées en 3 rangées. On en dénombre 12 500. Elles possèdent des cils disposés en W, encastrés dans une cuticule adhérant la membrane tectoriale.
Elles sont de différentes tailles tout au long de la cochlée : très courtes à la base (25 µm), et plus longues et lâches à l’apex (70 µm). Leur paroi latérale est constituée citernes latérales et de protéines membranaires (prestine), possédant des mitochondries, et ayant des propriétés contractiles ioniques (voltage-sensible).
Les mitochondries permettent un auto-contrôle de son gain, et jouent le rôle d’amplification cochléaire, mettant en mouvement la membrane tectoriale afin qu’elle incline les cils de la cellule ciliée interne correspondante. En bas de la cellule ciliée, on observe un compartiment synaptique renfermant les mitochondries.
Les cellules ciliées externes sont connectées à deux types de fibres nerveuses :
- Des fibres afférentes (bouton afférent spiral), ou terminaisons de neurones ganglionnaires de type II, qui permettent une connexion au système nerveux central, et qui pourraient être responsables d’un autocontrôle de l’électromobilité de la cellule ciliée.
- Des fibres efférentes, avec des larges terminaisons vésiculées (système efférent médian). Ces fibres efférentes innervent richement les cellules ciliées externes, et régulent leurs propriétés mécaniques.
Les cellules ciliées externes présentent deux temps de réponse :
- Une réponse rapide, de 20 à 25 kHz, voltage sensible, pendant laquelle la cellule ciliée suit les vibrations fréquentielles par modification de sa longueur grâce à la prestine (électromotilité).
- Une réponse lente, liée à la réponse du cytosquelette (actine et spectrine), permettant d’adapter la réponse de la cellule ciliée externe. Cette réponse est contrôlée par le système efférent médian de l’olive supérieure.
Les cellules ciliées externes jouent un rôle de régulation active de la sensation sonore en amplifiant les sons faibles et en amortissant les sons forts. Un traumatisme sonore peut endommager les cellules ciliées et provoquer une surdité irréversible ainsi qu’une hypersensibilité aux sons forts par effet de recrutement.
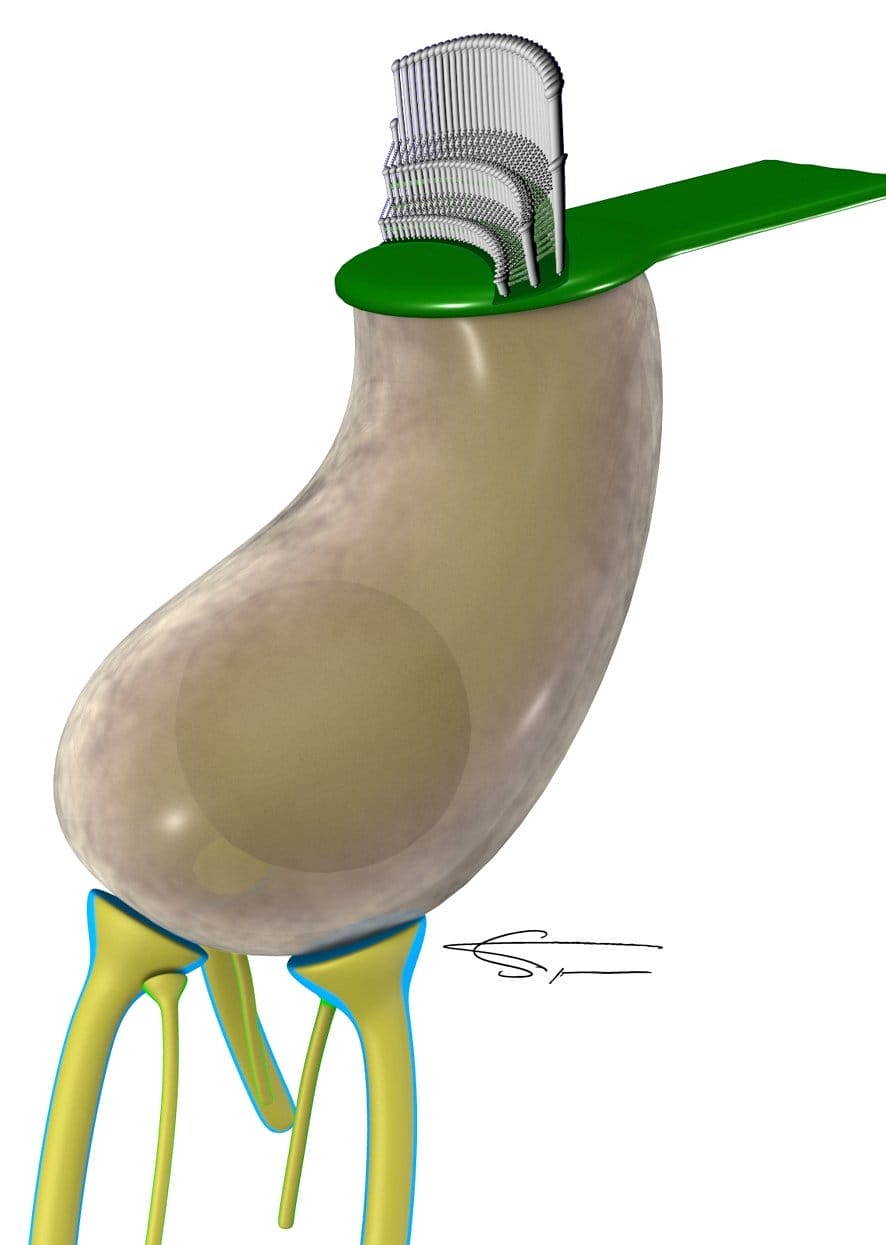
Les cellules ciliées internes
Les cellules ciliées internes (CCI), en forme poire, sont organisées en une rangée. On en dénombre 3500 environ.
Une cellule ciliée possède des cils libres, répartis le long d’une ligne, qui entrent en contact avec la membrane tectoriale (la rangée des cils les plus longs) lorsque les contractions des cellules ciliées externes le permettent.
Cela entraîne alors une déflection des cils et une dépolarisation de la cellule (par influx de cations K+). Cette dépolarisation entraîne, par une chaîne de réactions d’échanges de molécules, la libération d’un neuromédiateur : le glutamate.
Celui-ci stimule les extrémités dendritiques des fibres spirales efférentes du nerf auditif. Cette stimulation sera conduite jusqu’aux voies auditives centrales. Environ 10 à 20 fibres nerveuses sont en contact avec une cellule ciliée interne.
Seul un faible pourcentage de CCI ou de Fibres nerveuses auditives FNA sont nécessaires pour préserver un seuil tonal normal dans le silence, tant que les CCE restent intactes (Furman et al. 2013). L’oreille peut être endommagée et pour autant l’audiométrie tonale peut rester normale alors que nous avons la sensation de mal entendre notamment dans le bruit, il s’agit alors d’une surdité dite “cachée” ou Hidden Hearing Loss.
Traduction et codage dans l’oreille interne
La transduction mécano-électrique
La transduction mecano électrique peut être découpée en plusieurs étapes que nous décrivons ci-dessous :
- Le son a précédemment été transmis par le tympan et les osselets sous forme de vibration, mettant en vibration, via la fenêtre ovale, les liquides péri-lymphatiques de la rampe vestibulaire, puis, étroitement reliés, ceux de la rampe tympanique, jusqu’à la fenêtre ronde.
- Cela met en vibration la membrane basilaire (membrane basse du canal cochléaire), de haut en bas, et de manière plus importante sur une petite partie de la membrane en fonction de la fréquence du son (tonotopie passive).
- Les stéréocils des cellules ciliées externes correspondantes se retrouvent alors déplacés vers la membrane tectoriale, située juste au-dessus, auxquels ils sont implantés par la plaque cuticulaire. Ils sont alors basculés vers l’extérieur.
- Des canaux de la cellule ciliée externe, sensibles à l’étirement, s’ouvrent, et permettent l’entrée passive d’ions potassium K+, par un gradient d’ions positif de l’endolymphe (baignant dans le canal cochléaire et composé essentiellement d’ions K+ fournis par la strie vasculaire), vers le corps cellulaire de la cellule ciliée.
- Cette entrée de K+ entraîne la dépolarisation de la cellule ciliée externe. En réponse, elle se contracte par électromobilité, et permet une amplification de la vibration initiale ainsi qu’une augmentation de la sélectivité fréquentielle (tonotopie active).
- Les plus longs stéréocils de la cellule ciliée interne correspondante se retrouvent alors en contact avec la bande de Hensen de la membrane tectoriale. La cellule ciliée interne est alors dépolarisée par le même mécanisme de déflexion des stéréocils et entrées de K+ dans la cellule.
- Cela entraîne une entrée de calcium Ca2+ dans la cellule ciliée interne, et, par une chaîne de réactions, la libération, après migration des vésicules synaptiques, d’un neurotransmetteur dans la fente synaptique sous la cellule ciliée : le glutamate. Celui-ci se fixera sur un récepteur des terminaisons des fibres du nerf auditif, entraînant la création d’un potentiel d’action envoyé jusqu’au cerveau.
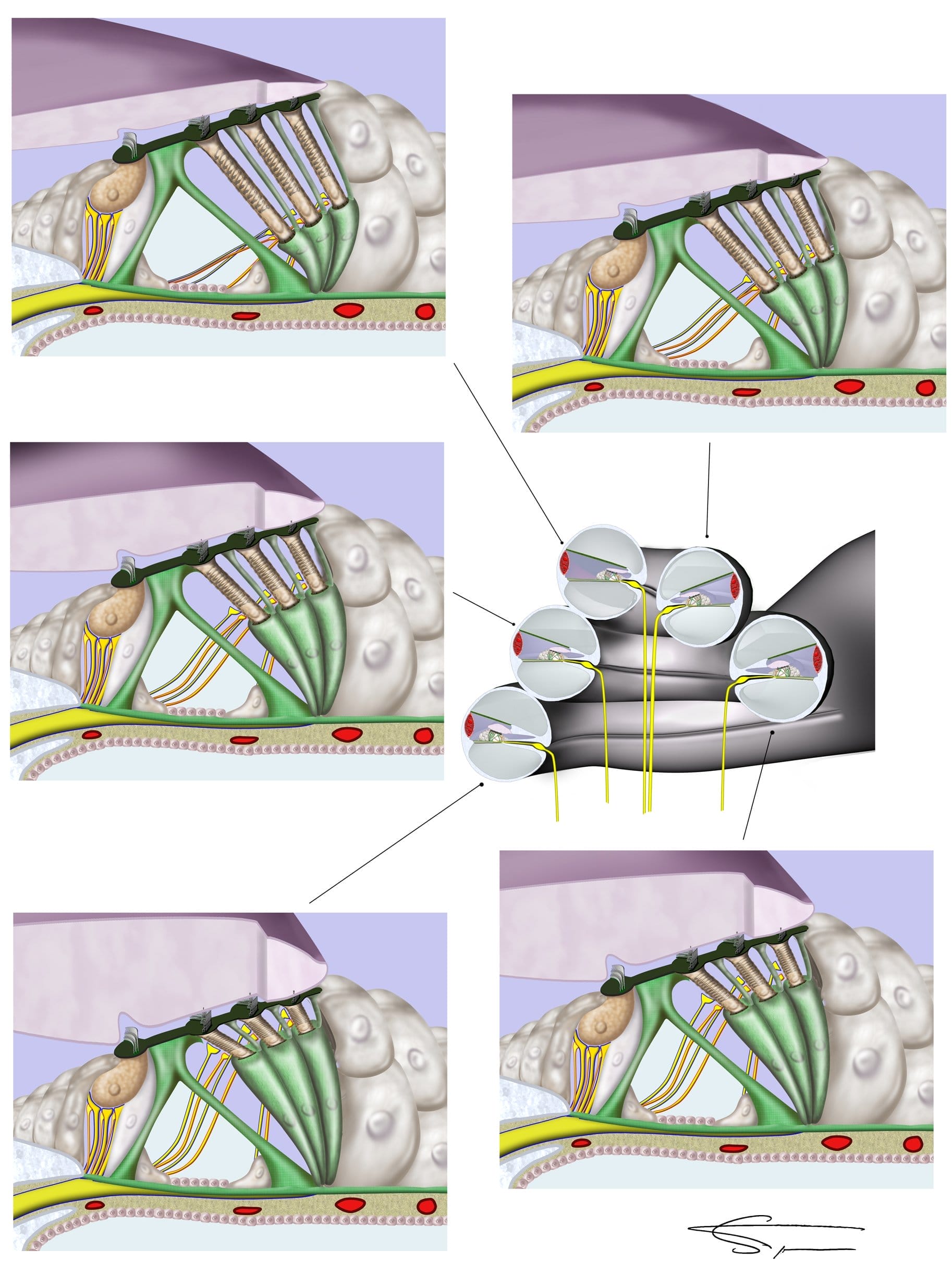
Le codage fréquentiel
La tonotopie passive:
La lame spirale, supportant le canal cochléaire, voit sa structure cochléaire évoluer tout au long du limaçon, de la base à l’apex. Cette différence de structure anatomique constitue la première sélectivité fréquentielle de la cochlée. La longueur de la membrane va effectivement s’accorder à la longueur d’onde des sons. La membrane basilaire vibrera ainsi plus fortement à un endroit précis de l’organe de corti, en fonction de la fréquence du son, en raison de cette différence anatomique de la base à l’apex. A la base, la membrane est courte et épaisse, et correspondra aux fréquences aigues, (jusque 20 kHz) tandis qu’à l’apex, la membrane est longue et fine, et correspondra aux sons graves (jusque 20 Hz).
La tonotopie active:
La sélectivité fréquentielle de la tonotopie passive est affinée par la tonotopie active, créée par la contraction des cellules ciliées externes (CCE) concernées (courbes d’accord). Cet accord en fréquence (tunning) permet un gain d’intensité de 50 dB, et permet d’entendre différemment deux sons de fréquence différente. Cette sélectivité sera conservée au niveau de la cellule ciliée interne et du nerf auditif.
La cellule ciliée externe peut également intervenir en cas de son trop fort, en inhibant légèrement la réponse. A forte intensité, la spécificité fréquentielle sera réduite par la colonisation des cellules ciliées voisines de celles de la stimulation. Un ajustement de la réponse des cellules ciliées externes sera également engendré par des fibres afférentes de centres protubérantiels répondant aux stimulations reçues depuis les cellules ciliées externes, et étant dépendants de chaque personne.
Le verrouillage de phase:
Lorsque les sons sont inférieurs à 3kHz, le codage en fréquence est également permis par le verrouillage en phase (phase locking) par la cellule ciliée interne. En effet, la cellule ciliée interne se dépolarise et se repolarise alternativement, en phase avec la fréquence de la stimulation sonore. Cela entraîne une libération de glutamate et donc la création d’un potentiel d’action en phase avec la stimulation sonore.
Ce verrouillage de phase est très efficace pour les fréquences graves (ex : 500 Hz), et son efficacité diminue vers 2000 Hz, car la CCI n’est plus assez rapide pour se dépolariser/repolariser en suivant la fréquence de stimulation. Au-delà de 3000 Hz, on observe une perte de synchronisation des potentiels d’action par rapport au cycle de la fréquence de stimulation. Seul la composante continue de la stimulation est présente, et la cellule ciliée interne ne code plus que l’enveloppe de la stimulation sonore.
Le codage de l’intensité
On peut tout d’abord l’observer par une augmentation du nombre de potentiels d’action des neurones auditifs. On sait aujourd’hui qu’il existe des fibres auditives répondant à différents niveaux sonores, en fonction de leur activité spontanée (activité existant en l’absence de stimulation sonore).
On peut observer trois différentes populations de fibres, qui se relaient sur toute la dynamique auditive (de 0 à 120 dB) :
- Les fibres à haute activité spontanée, codant les intensités faibles. Elles présentent des densités post-synaptiques étendues. Elles saturent rapidement.
- Les fibres à moyenne activité spontanée, codant les intensités moyennes, et prenant le relai quand les fibres très sensibles saturent.
- Les fibres à faible activité spontanée, codant les intensités fortes. Elles présentent des densités post-synaptiques de surface réduite.
La réponse d’une fibre peut être décrite par un graphe, montrant une augmentation de l’activité au début de la stimulation sonore (pic on) correspondant à la libération du glutamate, puis une diminution (adaptation) pour atteindre un plateau (taux constant de décharge), et une diminution importante à l’arrêt de la stimulation.
Le système nerveux auditif
Il est constitué de plusieurs relais neuronaux, ayant tous un rôle différent pour une intégration complète du message sonore. La tonotopie se conservera tout du long des voies auditives nerveuses.
La synapse
La synapse, située sous la cellule ciliée interne, est le lieu de libération du neurotransmetteur appelé glutamate. Il est le reflet du couplage entre cellule ciliée interne et terminaison des fibres du nerf auditif.
Les synapses des cellules ciliées sont dotées de rubans synaptiques (constitué essentiellement de protéines RIBEYE), une structure dense aux électrons. Ces corps synaptiques sont ancrés à la membrane plasmique de la CCI et entourés de vésicules synaptiques contenant du glutamate. On compte 10 à 20 rubans synaptiques par CCI, chaque zone étant connecté à la terminaison dendritique d’une seule fibre auditive. Il y a donc 10 à 20 neurones ganglionnaires de type I par CCI.
La libération du glutamate au sein de la synapse :
La stimulation sonore entraîne l’ouverture des canaux sensibles à l’étirement, et la dépolarisation de la CCI. Les canaux calciques sensibles au potentiel s’ouvrent, provoquant une entrée de calcium proche des rubans synaptiques. Le remplissage de glutamate dans les vésicules synaptiques est assuré par le transporteur VGLUT3, et l’otoferline serait le détecteur calcique de l’éxocytose. Le glutamate sera alors libéré dans la synapse, puis activera les récepteurs AMPA des fibres auditives afférentes, premier relai des voies auditives.
Les synapses en ruban sont particulièrement fragiles et l’exposition au bruit peut les endommager provoquant une gêne auditive dans le bruit.
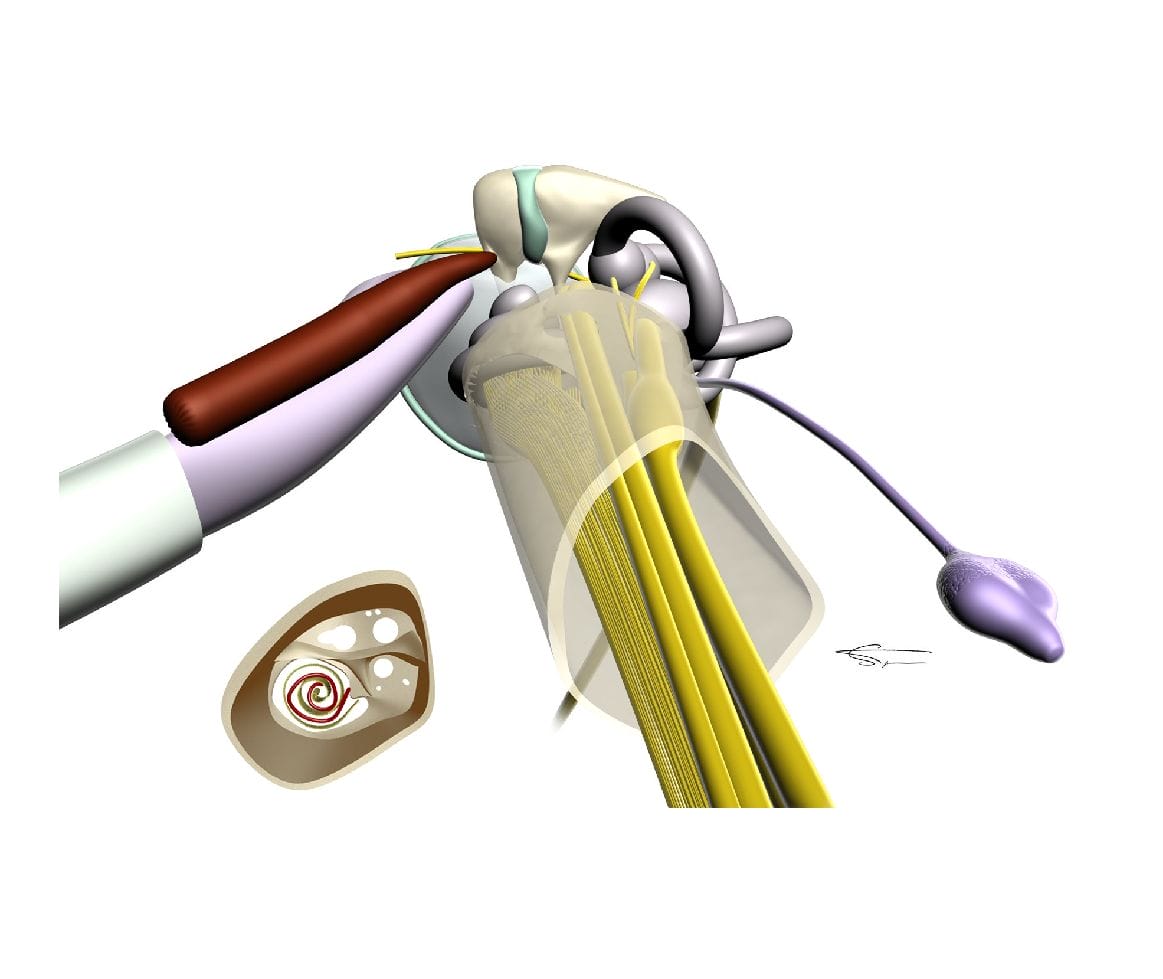
Le nerf auditif
Le conduit auditif interne est situé en dedans du labyrinthe osseux long de 8 à 10 mm et large de 4 à 5 mm. Il est divisé en 2 étages :
L’un supérieur où s’engage le nerf facial et la branche supérieure du nerf vestibulaire. L’étage inférieur où s’engage les rameaux du nerf cochléaire et la branche inférieure du nerf vestibulaire.
Le nerf auditif (VIII) est essentiellement sensoriel, il comprend le nerf cochléaire et le nerf vestibulaire.
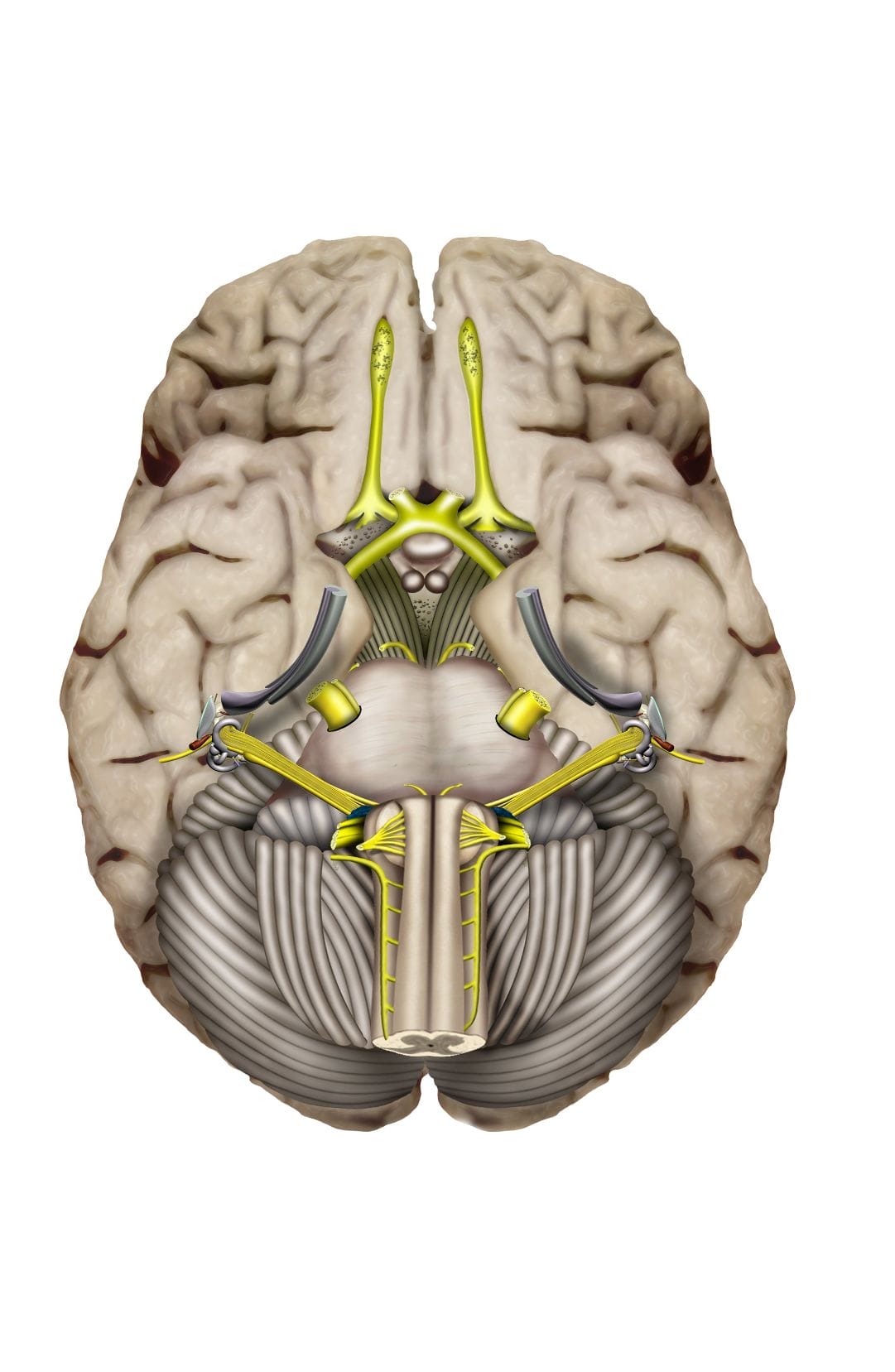
Le nerf cochléaire
Il naît de l’organe de Corti à l’intérieur de la cochlée membraneuse. Ses fibres nerveuses suivent d’abord les canaux de la lame spirale osseuse pour rejoindre leur corps cellulaire situé dans le canal spiral, au niveau du modiolus.
L’ensemble des corps cellulaires forme le ganglion de spiral de Corti. Celui-ci comprend environ 35 000 neurones sensoriels primaires (ou proto-neurones). Ces neurones ganglionnaires sont bipolaires, et peuvent être de type I et II qui permettent l’innervation afférentes des cellules ciliées. Ils se prolongent jusqu’aux noyaux cochléaires du tronc cérébral. Les fibres nerveuses suivent ensuite les canaux de la columelle, puis la fossette cochléaire située au fond du conduit auditif interne.
Le nerf cochléaire innerve les cellules ciliées de manière double : les informations sont véhiculées sous forme de boucle vers le système nerveux par les fibres afférentes, et inversement en provenance du système nerveux central par des fibres efférentes.
La CCI est innervée par le système afférent radial (neurones de type 1 du ganglion spiral), qui relie la cochlée aux noyaux cochléaires. Les neurones de type I sont myélinisés (95%) et de grosse taille, avec une branche périphérique unique connectée aux CCIs. Il y en a environ 10 neurones de type 1 par CCI. Elle est également innervée par le système efférent latéral, provenant de l’olive supérieure homolatérale, effectuant un rétrocontrôle sur la synapse, notamment pour éviter une excitotoxicité.
La CCE est innervée par le système afférent spiral (terminaisons dendritiques des neurones ganglionnaires de type 2 du ganglion spiral), dont le rôle est inconnu à ce jour. Les neurones de type II son non myélinisés et petits. Il y en a souvent un pour 10 CCEs. Elle est également innervée par le système efférent médian (terminaisons axoniques des neurones situées dans le complexe olivaire supérieur médian), permettant de réguler l’électromotilité des CCEs.
Le nerf auditif est constitué de différentes couches de fibres respectant la tonotopie de la cochlée : les fibres centrales viennent de l’apex de la cochlée, et sont entourées de fibres provenant plutôt de la base de la cochlée.
On distingue les voies auditives centrales primaires (dédiées à l’audition), et non primaires (convergence d’un ensemble d’informations). Leur fonctionnement simultané est nécessaire pour la perception consciente d’un son.
Les voies auditives primaires
Elles permettent de transporter l’information codée par la cochlée. Elles sont rapides (fibres myélinisées), commencent par le ganglion spiral de la cochlée (décrit précédemment) et se terminent sur le cortex auditif primaire. Elles présentent plusieurs relais effectuant chacun une tâche spécifique de décodage et d’interprétation.

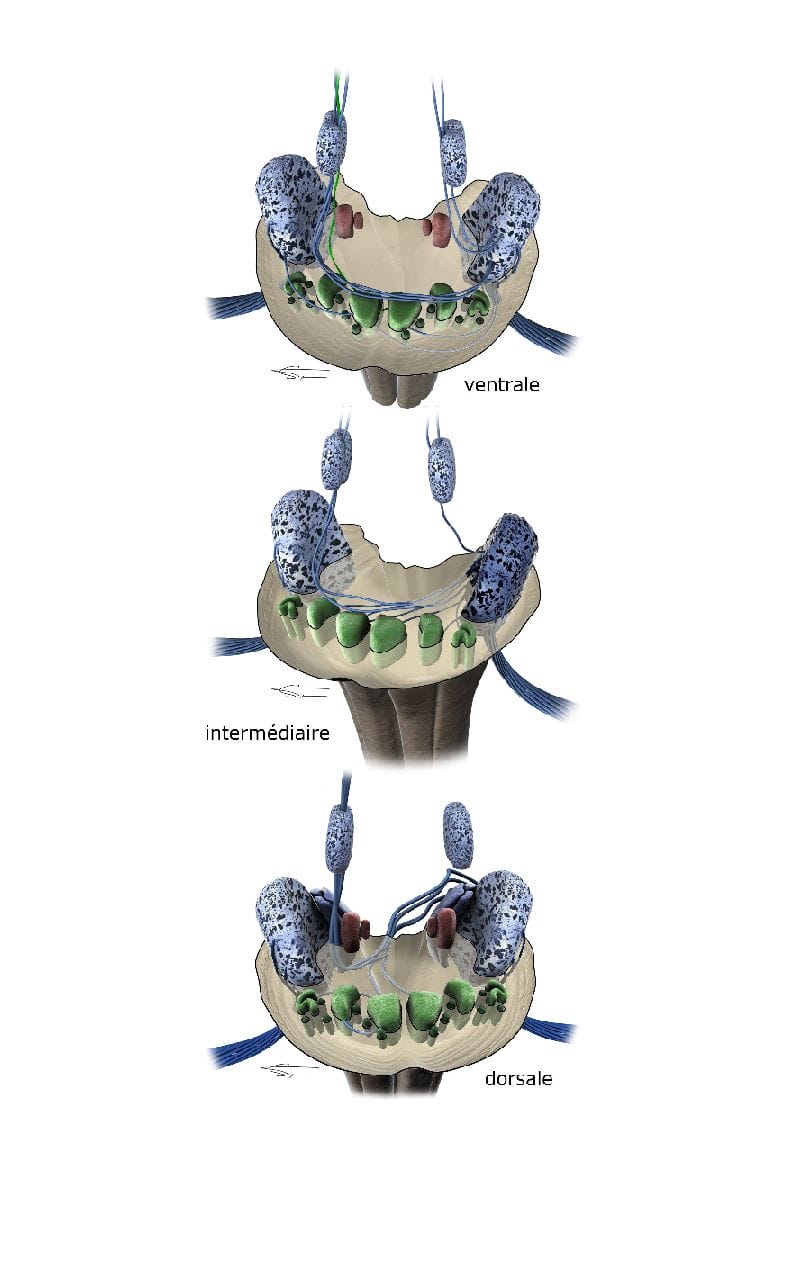
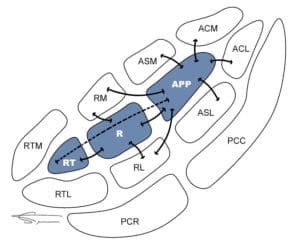
1er RELAIS
Le premier relai est constitué des noyaux cochléaires du tronc cérébral (au niveau bulbo pontique), recevant les axones des neurones de type I du ganglion spiral. Il permet de coder la durée (temps et début et fin du stimulus), intensité et fréquence du message. La tonotopie est effectivement conservée par le plan d’isofréquence.
Chaque neurone a une fonction spécifique : les neurones globulaires et sphériques codent la durée, les neurones octopus codent le début de la stimulation, et les neurones fusiformes et pyramidaux s’activent au début et à la fin du stimulus.
Les noyaux cochléaires permettent également d’activer ou d’inhiber les neurones les uns par rapport aux autres pour mieux analyser et répartir le signal auditif primaire pour les relais supérieurs.
Chaque fibre se divise en V et donne deux branches, l’une pour le noyau antérieur (ventral ascendant), et un autre pour les noyaux postérieur et dorsale (dorsale descendant).
Les neurones cochléaires projettent leur fibre vers d’autres noyaux par des stries acoustiques qui permettent de transporter l’information aux relais supérieurs. On distingue ainsi la strie dorsale (constituée des fibres des neurones pyramidaux et multipolaires), la strie intermédiaire ou noyau ventral postérieur (constitué des fibres des neurones octopus) et la strie ventrale ou corps trapézoïde (constitué des axones des neurones sphériques globulaires et multipolaires).
2éme RELAIS
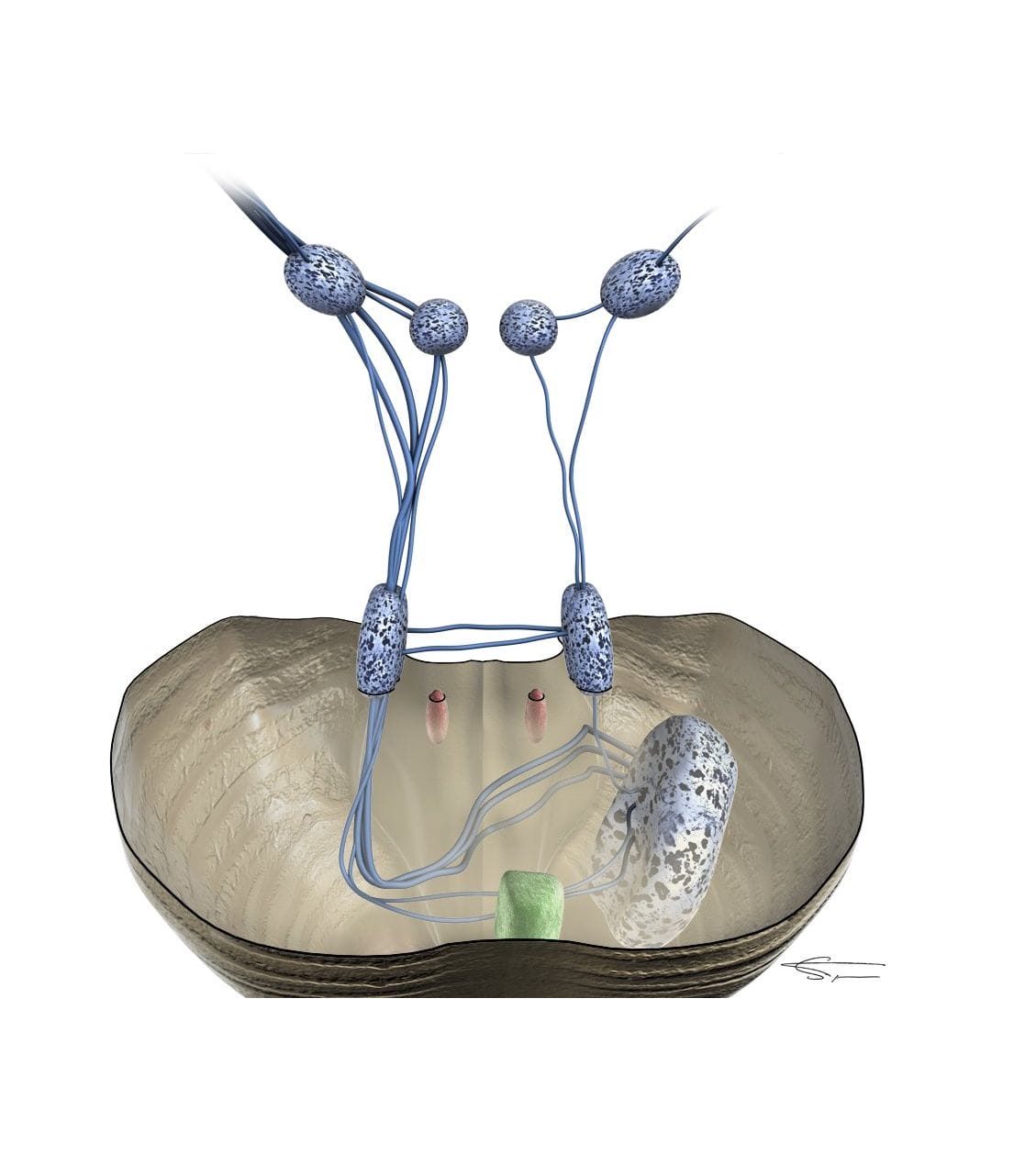
Le deuxième relai est le complexe olivaire supérieur qui reçoit les fibres auditives ayant croisées la ligne médiane.
Il est constitué de plusieurs noyaux avec des projections ascendantes (vers le cortex) et descendantes (vers la cochlée). Certaines fibres ascendantes font un premier relai dans le noyau du lemnisque latéral, d’autres vont directement au colliculus inférieur.
Il permet une analyse et un filtrage complexe de l’information sonore allant vers le cortex, mais aussi la création de reflexes et une adaptation protectrice du système auditif en cas de danger. Il est également très impliqué dans la localisation des sons.
L’activité du COS est contrôlée par différents relais auditifs (supérieurs, et noyaux cochléaires et formation réticulée), et il se projette sur de nombreuses structures :
- Formation réticulée (réactions végétatives, alerte, éveil cortical…)
- Noyaux cochléaires, pour un rétrocontrôle
- Cochlée (systèmes efférents latéral et médian), pour un rétrocontrôle. Le système efférent latéral permet notamment de protéger la synapse entre la CCI et la fibre du nerf auditif d’une excitotoxicité (traumatisme ou ischémie) par libération de la dopamine. Le système efférent médian permet, par les terminaisons axoniques des neurones du complexe olivaire supérieur médian, de modérer l’électromotilité des CCEs.
- Noyaux du nerf facial (réflexe stapédien) et du nerf trijumeau (réflexe du marteau), pour une protection des structures de l’oreille interne contre les sons forts (> 80 dB). Il fonctionne par activation, après une boucle reflexe, des muscles reliés au stapédien et au marteau dans l’oreille moyenne afin de rigidifier la chaîne ossiculaire et de limiter la propagation de la vibration sonore. Ce reflexe n’est cependant fonctionnel qu’en dessous de 2000 Hz, est fatigable, et trop lent (30 ms) pour des bruits impulsifs.
- Noyaux du nerf oculomoteur latéral, pour visualiser la source sonore.
La tonotopie est conservée à ce niveau (fréquences graves dans les aires dorsales, fréquences aigues dans les aires ventrales), et correctement transmise aux relais supérieurs.
3éme RELAIS
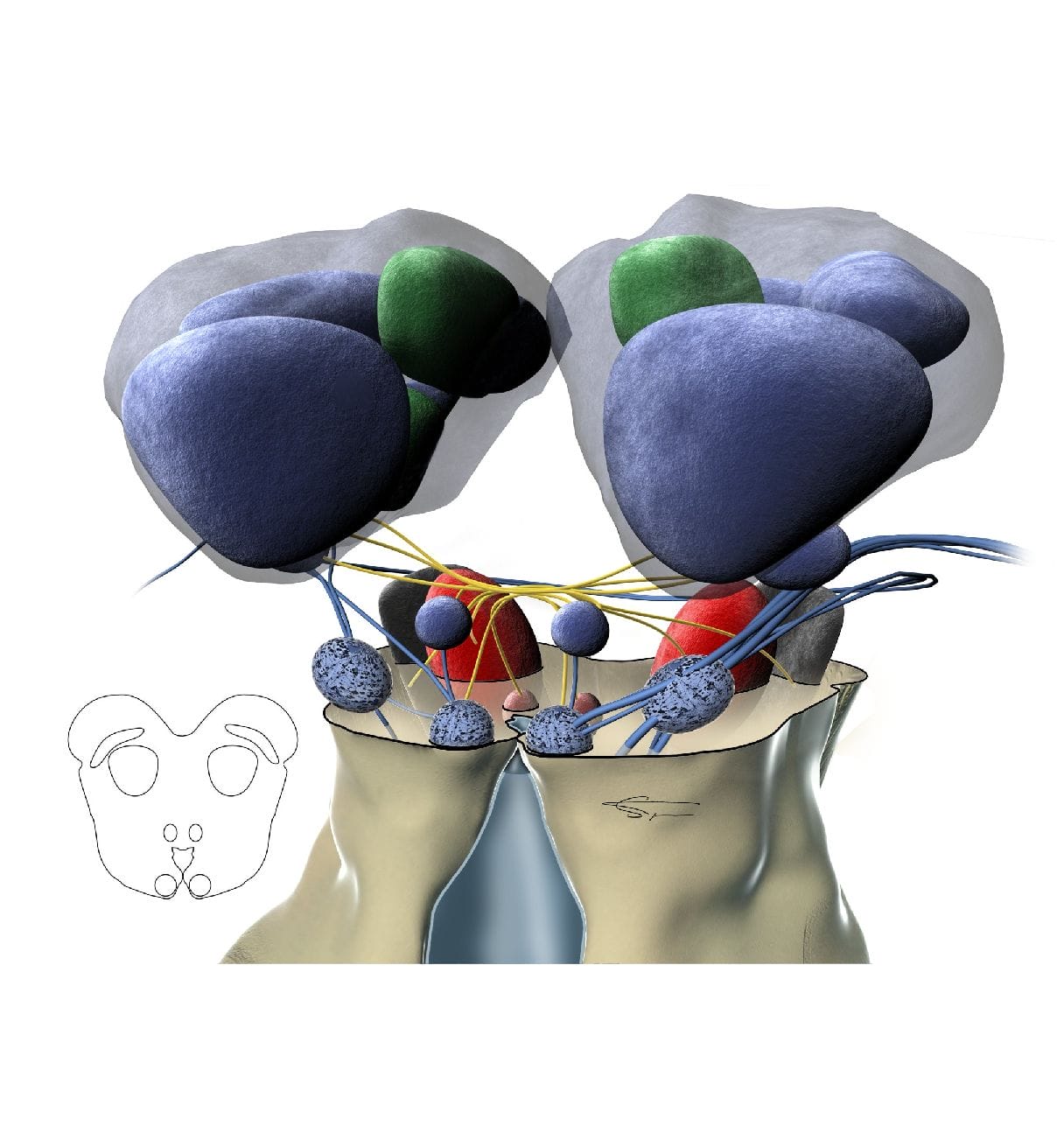
Les fibres montent par le faisceau du lemnisque latéral au colliculus inférieur (mésencéphale). Le noyau du lemnisque latéral est constitué de groupes de neurones disposés en un complexe ventral (composé de neurones monoauraux impliqués dans l’analyse de la durée d’un son complexe), et un noyau dorsal (composé de neurones binauraux recevant l’information des deux oreilles et jouant dans la localisation des sons).
Le colliculus inférieur, dans le mésencéphale, est composé du cortex externe et dorsal (impliqué dans l’analyse du langage et des nouveaux sons), et d’un noyau central. Ce dernier présente une organisation tonotopique complexe, recevant les basses fréquences de l’oreille ipsilatérale, et les hautes fréquences de l’oreille controlatérale. Il permet aussi un décodage des différences interaurales d’intensité et de temps. Le Colliculus inférieur est également impliqué dans la localisation des sources sonores (surtout dans le plan horizontal et vertical). Son activité est contrôlée par le cortex et le thalamus.

4éme RELAIS
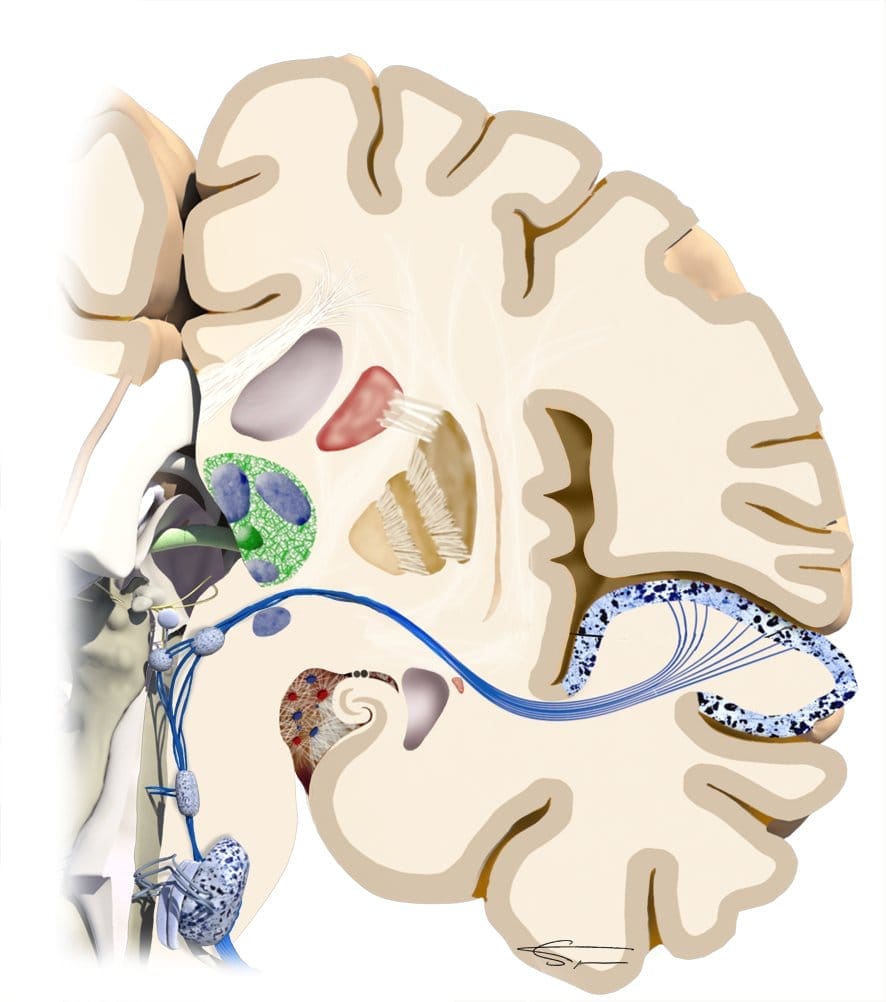
Le 4ème relai est situé dans le corps genouillé médian (thalamus), permettant l’intégration du son (préparation de la réponse motrice).
5éme RELAIS
Le 5ème relai est un neurone permettant de relier le thalamus au cortex auditif. L’information sonore est alors reconnue, et mémorisée, pour création d’une réponse volontaire.
Les voies auditives non primaires
Elles apparaissent juste après les premiers relais (noyaux cochléaires), par l’intermédiaire de fibres qui s’insèrent dans la voie réticulaire ascendante communes à toutes les fonctions sensorielles.
Les relais de la voie non primaire sont notamment la formation réticulée, le thalamus non spécifique, et enfin le cortex poly-sensoriel. Ils jouent le rôle de filtrage des sons (quelles informations sonores sont prioritaires). Ils sont en lien étroit avec les centres d’éveil et de motivation, et les centres de la vie végétative. Ils sont donc aussi reliés avec l’hypothalamus et les centres végétatifs.
Le vestibule
Le labyrinthe osseux de l’oreille interne contient le vestibule, qui est le siège de l’équilibre.
Il est divisé en deux parties : le système canalaire (canaux semi circulaires) qui répond aux mouvements de la tête, et le système otolithique (utricule et saccule) qui répond aux accélérations linéaires (horizontales ou verticales).
Le vestibule fait partie d’un ensemble d’organes (yeux, récepteurs proprioceptifs, muscles…) reliés au système nerveux central sous forme de boucle rétroactive, qui régissent l’équilibre du corps humain et la coordination des mouvements.
Une atteinte du système vestibulaire peut entraîner des vertiges et vomissements, ainsi qu’une sensation d’instabilité.
Le système canalaire
Il est formé de trois canaux semi-circulaires (CSC) membraneux. Ce sont des tubes qui ont la forme d’une boucle incomplète, implantés sur les parois de l’utricule, et qui ont une disposition orthogonale. Ils réagissent aux mouvements de la tête.
- Le CSC supérieur ou antérieur dans un plan vertical faisant un angle de 45 ° avec le plan sagittal du corps, ouvert vers l’avant.
- Le CSC postérieur dans un plan vertical faisant un angle de 45 ° avec le plan sagittal du corps, ouvert vers l’arrière.
- Le CSC externe ou horizontal dans un plan horizontal.
Les CSC présentent une extrémité ampullaire, appelée ampoule qui renferme la zone sensorielle qui se nomme la crête ampullaire. Les branches non ampullaires des canaux postérieurs et supérieurs se réunissent en un canal commun appelé crus commune.
La crête ampullaire est formée de cellules de soutien, et de cellules sensorielles (cellules ciliées) comportant une touffe ciliaire de stéréocils, de taille décroissante depuis le kinocil. Ces cellules sensorielles sont de 2 types, I et II, ayant une forme, une connexion synaptique et une fonction spécifique. Les cils de ces cellules sont au contact de la cupule (matrice protéique, structure gélatineuse) de même densité que l’endolymphe. La cupule obstrue complètement la lumière de l’ampoule. La touffe ciliaire des cellules sensorielles constitue une véritable antenne de réception des accélérations subies par la tête.
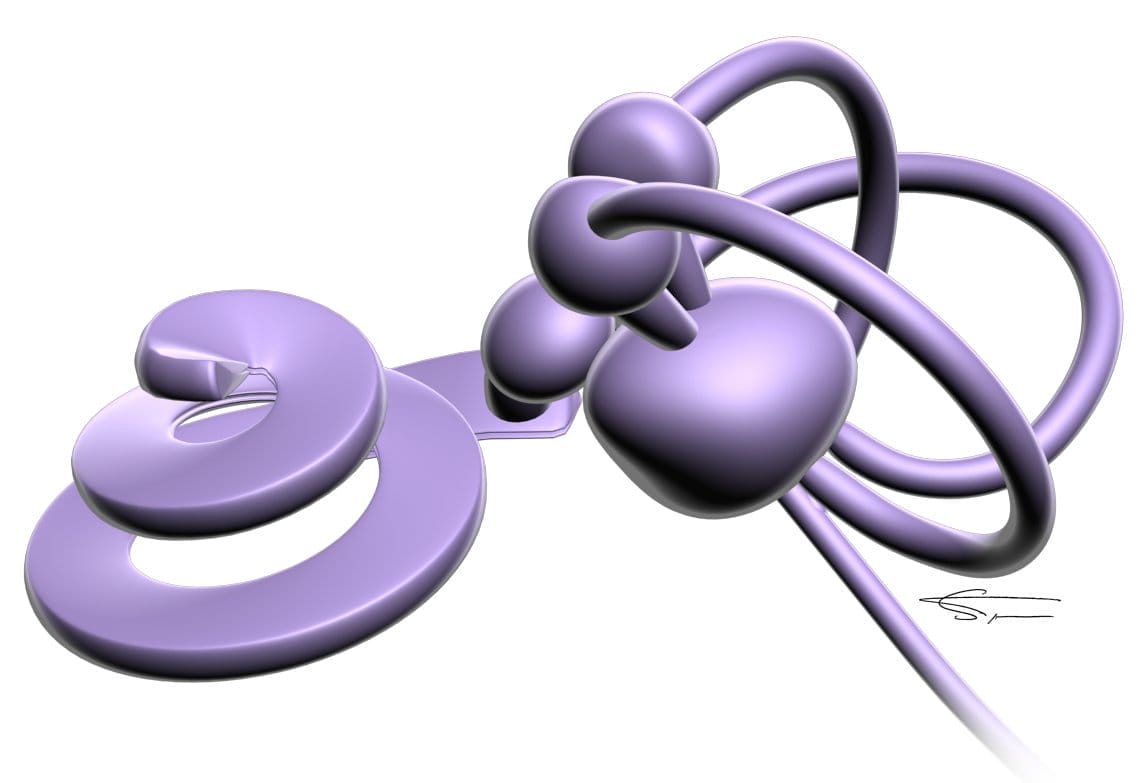
Les organes otolithiques
L’utricule et le saccule sont le siège des macules otolithiques utriculaires et sacculaires disposées perpendiculairement l’une par rapport à l’autre. Ils réagissent aux déplacements horizontaux et verticaux.
- La macule utriculaire est horizontale (dans le même plan que le canal semi-circulaire externe soit 30° par rapport au plan horizontal). Elle est donc concernée par les déplacements linéaires dans le plan horizontal.
- La macule sacculaire est placée dans un plan sagittal grossièrement perpendiculaire à la précédente. Elle est donc concernée par les déplacements linéaires dans le plan vertical.
Les macules sont constituées de trois couches :
- Une couche cellulaire, faite de cellules de soutien sur lesquelles reposent des cellules ciliées sensorielles de type I et II. Ces cellules émettent à leur face supérieure des prolongements appelés stéréocils, disposés en rangées de hauteur croissante avec un kinocil surplombant la plus haute.
- Une couche gélatineuse, ou membrane otoconiale, engluant l’extrémité de la plus haute rangée de stéréocils et le kinocil.
- Une couche otoconiale, ou otolithes, cristaux de carbonate de calcium, alourdissant la membrane otoconiale, la rendant hyperdense par rapport à l’endolymphe.
Chaque cellule ciliée a une polarisation fonctionnelle, définie par la position du kinocil par rapport à la touffe stéréociliaire. Seul un déplacement mécanique selon cet axe est susceptible de stimuler ou d’inhiber la cellule. Un mouvement dirigé vers le kinocil produit une excitation, tandis qu’un mouvement en sens opposé produit une inhibition. L’orientation fonctionnelle de chaque cellule ciliée varie de part et d’autre d’une dépression médiane, la striola qui divise la macule en 2 parties.
- Dans la macule sacculaire, les cellules sensorielles se tournent le dos, c’est à dire que les kinocils sont orientés vers l’extérieur (les vecteurs d’orientation sont striolafuges).
- Dans la macule utriculaire, les cellules sensorielles se disposent en vis-à-vis, c’est-à-dire qu’elles se font face, les kinocils sont orientés vers la striola (les vecteurs d’orientation sont striolapètes).
Le nerf vestibulaire
C’est le 1er neurone des voies vestibulaires, il est formé de cellules en T, analogues aux neurones des nerfs spinaux. Il prend son origine au fond du conduit auditif interne par la confluence du nerf vestibulaire supérieur, résultant de la fusion du nerf utriculaire et du nerf des CSC supérieur et externe, ainsi que du nerf vestibulaire inférieur relié au saccule et au CSC postérieur.
On distingue deux populations de neurones vestibulaires afférents :
- Les phasiques : spécialisés dans les mouvements rapides de hautes fréquences (connectés avec les cellules de type I).
- Les toniques : qui répondent aux mouvements lents de basses fréquences et dont la réponse calorique est la forme extrême (connectés avec les cellules de type II).
Il existe aussi des fibres nerveuses efférentes, dont l’origine se situe au niveau des noyaux vestibulaires et dont le rôle est de contrôler le système afférent et dont le neurotransmetteur est l’acétylcholine.
Le nerf vestibulaire a un trajet totalement intracrânien depuis les récepteurs jusqu’aux noyaux vestibulaires. Il est accolé, sur la presque totalité de son parcours, au nerf de la cochlée constituant la 8ème paire crânienne. Il chemine également avec le nerf facial. Les fibres nerveuses vestibulaires se rejoignent au niveau du ganglion vestibulaire qui est un renflement du nerf vestibulaire dû à la présence des corps cellulaires des neurones. C’est le ganglion de Scarpa.
Puis le nerf vestibulaire chemine dans le conduit auditif interne pour gagner l’angle pontocérébelleux et entrer dans le sillon bulbo-protubérantiel pour gagner les noyaux vestibulaires.